Ángela Baeza-Serrano*![]() | Nuria Oliver
| Nuria Oliver![]() | Feliu Sempere
| Feliu Sempere![]() | Tatiana Montoya
| Tatiana Montoya![]()
© 2023 IIETA. This article is published by IIETA and is licensed under the CC BY 4.0 license (http://creativecommons.org/licenses/by/4.0/).
OPEN ACCESS
Stable and active microbiological community of the activated sludge is essential for waste water treatment plants (WWTPs) to treat wastewater. The microbial community of activated sludge could be affected by the uncontrolled industrial discharges containing excess organic load with low nutrients or toxic compounds, consequently, it may have a negative impact in the purification of wastewater in WWTP. The effects of the industrial pollutants on biological treatment include: The inhibition of bacteria that biodegrade organic matter and remove nutrients; the reduction of the efficiency of solids separation and deterioration of settling properties of the flocs; and, eventually, a discharge of unfavorable effluent to the receiving environment. In this research, metabarcoding technology was used to elucidate the effect of industrial components on microbial communities in four plants treating municipal that were exposed to different spills: tannery, cannery, textile and fruit products industries.
activated sludge, industrial discharges, bacterial community, metabarcoding, tannery, cannery, textile, fruit products industry
Activated sludge is the most widely used secondary wastewater treatment system in the European Union (EU). It involves a wide variety of micro-organisms that biodegrade organic matter via oxygen oxidation and remove nutrients. The bacterial communities in activated sludge systems are diverse, complex, and dynamic and in addition, hence the influence on the operation and performance of the biological reactor [1].
The processes involved in wastewater treatment are determined by numerous factors such as influent characteristics, effluent requirements, WWTP size, local temperature, land-use fees, or geology, among others. Operating parameters of wastewater treatment influence the formation of complex microbial structures and microbial composition, which perform different metabolic pathways in the system and, thus, treated wastewater quality [2]. According to the study of Zhang et al. [3], influent composition is the primary determinant of the composition of bacterial communities in WWTPs. In the light of the above, it is of interest to study the effect of industrial discharges to WWTP on the bacterial community in activated sludge in urban WWTP system.
WWTPs, as biological systems, are susceptible to be negatively impacted by toxic compounds in influent wastewater which could disrupt the operation of WWTPs, especially regarding to its nitrifying capacity [4]. Such dysfunctions usually come from uncontrolled industrial discharges with excess toxic loads, which, futhermore, can be seasonal.
Industrial wastewaters are highly variable in terms of flow and composition. Their characteristics is not only different from one to another, but also various even in the same industry. For instance, the treatment of wastewater resulting from general fruit handling is conducted on the basis of disinfectants, detergents, fungicides and/or water-based waxy coatings used, and these components individually and/or in combination contribute to high ecotoxicity of processed wastewater from citrus warehouses. According to cleaning or peeling processes, not only is the wastewater of cannery industry mainly composed of organic matter and suspended solids, but it also has sometimes high conductivity with high or low pH. Tanning process uses acids, alkaline substances, chromium salts, solvents, tannins, sulphides, dyes, auxiliaries, and many others components; and these chemicals remain in the effluent [5]. Textile dyeing processes produce coloured wastewaters, with extreme pH, high organic matter (COD and BOD5) and different salts, surfactants, heavy metals, mineral oils as well as others toxic compounds [6].
In this study, the effects of different industrial discharges on activated sludge communities of four WWTP were characterized in order to address the impact of industrial discharge to WWTPs.
The activated sludge communities into the biological reactor of four WWTP located in central and eastern Spain were studied during a whole year. All WWTPs treated municipal wastewater but frequently received discharges from different types of industries. The design features of the WWTPs and their sources of industrial discharges are shown in Table 1.
In 2018, monthly samples of 5 mL activated sludge were taken from these WWTPs, and then frozen and transported on dry ice to laboratory for the metabarcoding analysis. In April, 2018, WWTP 4 had to empty the biological reactor due to maintenance operations so samples were not taken.
The extraction of genomic DNA of 2 mL of samples was conducted with Qiamp Power Fecal Mini kit (Qiagen) using enzymatic lysis and mechanic disruption in ADM-Lifesequencing S.L. laboratory (Valencia, Spain). A total of 50 ng of DNA was amplified following the 16S Metagenomic Sequencing Library Illumina 15044223 B protocol (ILLUMINA). In summary, primers were designed containing 16S rRNA gene universal primers and a universal linker sequence allowing amplicons for incorporation indexes and sequencing primers by Nextera XT Index kit (Illumina) [7].
Table 1. Characteristics of WWTPs and major industrial spills received
|
|
WWTP1 |
WWTP2 |
WWTP3 |
WWTP4 |
|
Treatment System |
Extended aeration |
Extended aeration |
Extended aeration |
IFAS |
|
Design Flow Rate (m3/d) |
14400 |
6114 |
6000 |
600 |
|
Population Equivalent |
91200 |
15375 |
40848 |
1445 |
|
Sector |
Citrus grocery stores |
Textile |
Tanneries |
Canneries |
|
Industrial Spills |
Fungicides, pesticides, waxes |
Metals, pH, chlorides, sulphates and dyes |
Acids, alkalis, sulphides, chromium |
Conductivity, pH, organic matter |
|
Reactor T ºC (max/min) |
31.8/7.4 |
25/17 |
27.6/10.2 |
29.3/17.4 |
|
Location |
La Pobla de Vallbona (Valencia) |
Bétera (Valencia) |
Canals (Valencia) |
Binaced (Huesca) |
Amplification indexes were introduced in the last analysis. 16S based libraries were quantified by fluorimetry using Quant‐iT™ PicoGreen™ dsDNA Assay Kit (Thermofisher). These were sequenced and previously pooled on the MiSeq platform (Illumina), with 300 cycles paired reads configuration. Bioanalyzer 2100 (Agilent) was used in order to assess the size of the pool and its amount with the Library Quantification Kit for Illumina (Kapa Biosciences). PhiX Control library (v3) (Illumina) was combined with the amplicon library (expected at 20%).
Sequencing data were obtained within approximately 56 h. The MiSeq instrument (MiSeq Control Software (MCS v3.1) was used for image analysis, base calling and data quality assessment.
Raw sequences, both forward and reverse, were merged in order to obtain the complete sequence using the BBMerge package of BBMap V.38 software. With such the used to trim the amplification primers from the sequences obtained in the sequencing phase in order to decrease the bias in the annotation phase.
Sequences lower than 200 nts were removed from the assay after the primers’ remotion because the shorter the sequences are, the greater the probability of generating short erroneous taxonomical group associations. Once the clean complete sequences were obtained, a quality filter was applied to them to delete poor quality sequences. Bases in extreme positions not reaching Q20 (99% well incorporated base in the sequencing phase) or a greater pared score, were removed. Later, sequences with average quality less than the Q20 threshold were deleted as well. FASTQ files were converted to FASTA files and CD-hit program version 4.8.1 was used with the aim of remove chimeras. NCBI 16S rRNA database was used to BLAST [8] the FASTA files without chimeras, using blastn version 2.2.29+. A python script developed by ADM-Lifesequencing S.L. for the annotation of each sequence at different phylogenetic levels was used in order to process the resulting XML files. Shannon and Chao Biodiversity indexes were calculated with Specaccum program implemented for R version 3.2.3.
3.1 Microbial communities diversity
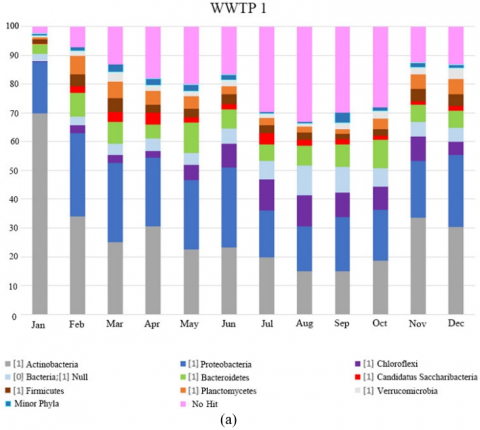
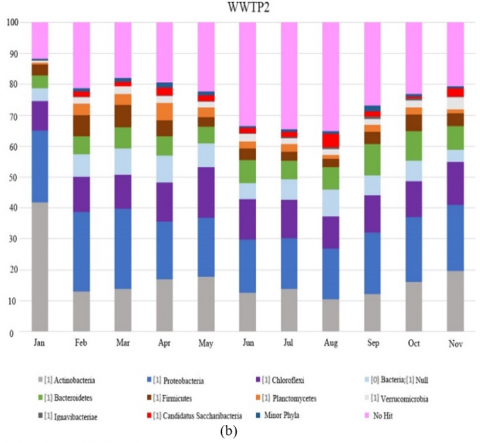
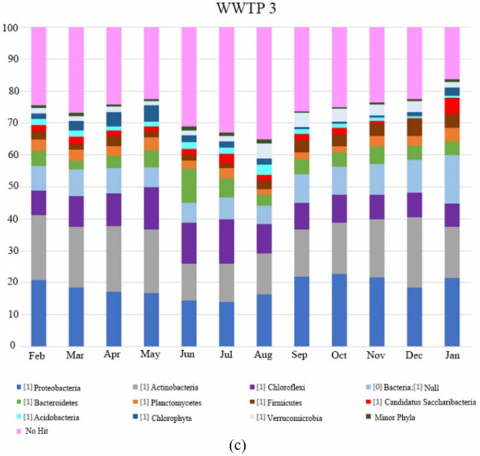
The diversity of microbial communities at phylum level in the biological reactors of the WWTPs studied over one year period is presented in Figure 1. Taxa represented in Figure 1 had relative abundance at a greater than 1% in at least one sample. The abundance was presented in terms of percentage based on the total number of bacterial sequences determined in each sample. Taxonomical identification was at a confidence threshold of 99%.
Proteobacteria phylum dominated most of the samples of WWTP 3 and 4 coinciding with the results obtained by Wang et al. [9] in 14 WWTPs located in China and with those found by Hu et al. [10] in 12 plants also in China. Wang et al. [11] described how the activated sludge population shifted in bulking periods from Proteobacteria to Actinobacteria dominated bacterial communities. The same situation was observed at WWTP 1 and 2. Almost the 70% of relative abundance in WWTP 1 and 42% in WWTP 2 are represented from Actinobacteria phylum coinciding with a bulking process caused by Candidatus Microthrix parvicella morphotype in January. In fact, almost 65% of the relative abundance at genus level in WWTP 1 and almost 7% in WWTP 2 were represented by the genus Candidatus Microthrix. After that event the dominant phylum changed to Proteobacteria.
Figure 1. Abundance of OTUs at phylum level (%) at (a) WWTP 1, (b) WWTP 2, (c) WWTP 3, and (d) WWTP 4
The phylum Firmicutes dominated in WWTP 4 in August 2018, associating with the beginning of peach canning campaign and concretely, with two spills detected in the inflow of the WWTP with high pH (which increased from an average value of 8.1 in July to a pH point value of 11.29 during pills) and changes in conductivity from a mean of 1,526 µS/cm in July to point values greater than 10,000 µS/cm during spills. Firmicutes are well represented in anaerobic sludge treatment systems and they are versatile in degrading a big array of environmental substrates [12].
The calculated species richness based on the Chao1 and Shannon index were used in this study. The Shannon Index values of activated sludge samples were similar among the four different plants, especially at high range (Table 2). The results indicated that the activated sludge in the WWTPs consisted of highly diverse microbial ecosystems. The results are different from those reported by Gonzalez-Martínez et al. [13], where they observed lower Shannon index and higher Chao 1 values in 10 different wastewater treatment systems located in Spain and Netherlands (Chao 1 1395,003-441,150 and Shannon index 5.137-2.831). The observation could be associated with the presence of industrial spills in wastewaters.
Table 2. Chao 1 and Shannon diversity indexes
|
Plant |
Shannon Value |
Chao 1 |
||
|
|
Max |
Min |
Max |
Min |
|
WWTP 1 |
5.017 |
3.572 |
2020 |
1569 |
|
WWTP 2 |
5.319 |
4.666 |
2349 |
1703 |
|
WWTP 3 |
5.435 |
5.299 |
2647 |
2001 |
|
WWTP 4 |
5.215 |
4.775 |
2496 |
1572 |
Actinomycetes, mainly Streptomyces species, are able to decolorize and mineralize textile dyes, and have been proposed in biodegradation treatments [6]. In WWTP 2 which received textile discharges, a greater relative abundance of this genus was detected up to 6.63%, while in the other plants without textile wastewater, the relative abundance was lower than 0.36%, indicating that textile wastewater did contribute to the abundance of Actinomycetes.
3.2 Population structure of nitrifying bacteria
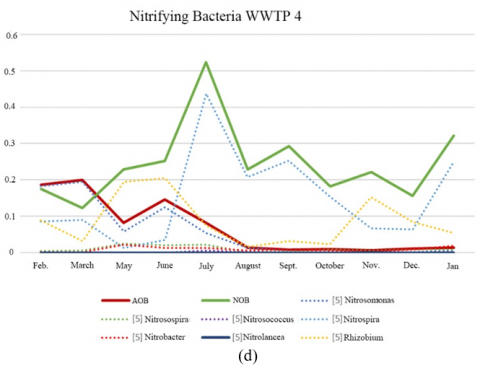
The most important genera of ammonia oxidizing bacteria (AOB) in WWTPs are Nitrosomonas and Nitrosospira [14, 15]. Nitrosomonas were the most abundant AOB in the samples from all plants, accounting up to 0.412% of the total sequences evaluated, indicating proper nitrification performance in the four WWTPs. Under poor nitrification conditions, low values for nitrifying bacteria were observed [16]. Nitrosospira was rarely detected in WWTP 1 but it was present in all samples from WWTP 4. Both genera are responsible for the nitrification of ammonium to nitrate.
The most diverse population of nitrifying bacteria was detected in WWTP 4, which received discharges from the cannery industry. This may be because it has an IFAS system that would facilitate the growth of nitrifying bacteria due to that biomass had possibility to grow on the carriers with a longer retention time.
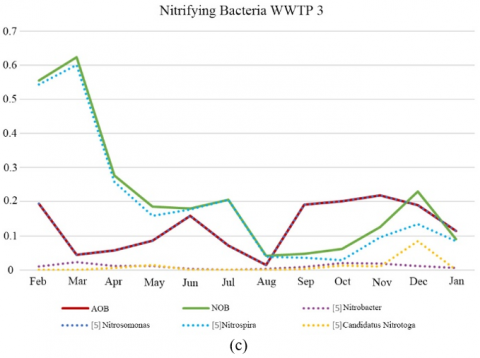
Lücker et al. [17] demonstrate that Nitrotoga was a functionally important nitrite oxidizers genera in 21 full-scale WWTPs located in Germany and Switzerland, where such genera often coexisted with Nitrospira but occasionally represented the only known NOB populations. In this study, Candidatus Nitrotoga was detected in WWTP 1, WWTP 2 and WWTP 3, being dominant against Nitrospira in WWTP 1 and, during autumn in WWTP 2 (Figure 2 (a), (b) (c) and (d)).
Figure 2. Relative abundance of nitrifying bacteria OTUs (%). (a) WWTP 1; (b) WWTP 2, (c) WWTP 3, (d) WWTP 4
According to Yao and Peng [18], a high a percentage of NOB in a nitrifying community indicates that the growth of NOB does not rely only on the nitrite provided by AOB, but also in nitrite provided by nitrate reducing bacteria, which could grow in the anoxic zones created inside the flocs. This could explain the fact that in WWTP 2, during August and September, although AOB accounted 0%, a high relative abundance of NOB was detected (Figure 2 (b)) without affecting nitrogen removal efficiency (RE) whose average was 84.67%.
The citrus season affecting WWTP 1 which started in September and lasted until March, 2018. Since September, a drop of AOB relative abundance but not NOB was observed in WWTP 1 (Figure 2 (a)). A drop in the nitrogen removal performance was also observed in the middle of September, and also in December and January when the AOB and NOB relative abundance are lower.
During April and May, no AOB and NOB was detected in WWTP 2 (Figure 2 (b)). This was associated with numerous spills with high conductivity, and containing chlorides and sulphites. Interestingly, nitrogen removal performance was virtually unaffected in the absence of nitrifying bacteria (average nitrogen RE 84.67%), which suggested the presence of other unknown genera of nitrifying bacteria.
WWTP 3 was affected by tanneries, especially in the run-up to the August and Christmas holiday periods due to cleaning of facility before it ceased to operate. In Figure 2 (c) a decrease in relative abundance of AOB and NOB was observed in August. Despite this, the population of nitrifiers was maintained, albeit at lower abundance, and nitrogen removal performance was not substantially affected (average nitrogen RE 89%).
WWTP 4 suffered two spills with high pH (>10) on 4th July and with high conductivity on 18th July (>10,000 µS/cm). An increase in relative abundance AOB and NOB was observed in July, with a subsequent decline in relative abundance of both bacterial groups as nitrogen removal efficiency decreased (from 90% up to 0%). High temperature and long sludge age could explain the increase in relative abundance of AOB and NOB in the months of May and June [19].
3.3 Evolution of microorganisms causing bulking and/or foaming
One of the most common operational problems in activated sludge system is sludge bulking, i.e., the massive proliferation of filamentous bacteria which leads to problems of sludge separation.
Candidatus Microthrix is a major cause of bulking and foaming. This morphotype has been described like fat, oil, and grease (long chain fatty acids) that consumers grow under critical conditions such as low temperatures and dissolved oxygen levels [20]. It is common to find problems caused by Ca. Microthrix during the winter season [21].
Ca. Microthrix genus was found in three of the four WWTPs. WWTP 1 had the highest relative abundance of this genus at almost 65% during a bulking episode that started in January and lasted over winter season. It decreased as temperature became higher in May, and its relative abundance increased again in September, coinciding with the beginning of the citrus season. In WWTP 2, high relative abundances were detected between January and May associated with low temperature with the abundance between 2.8% and 6.9%. On the other hand, high relative abundance was detected in a very localized manner in WWTP 3 during April and May (4.2% and 16.6% respectively) and occurred after December (18.8%), as mentioned above due to the cleaning facility prior to the Easter and Christmas holidays of the tanning industries which discharged wastewater, and affecting this WWTP. During the rest of the year, its relative abundance was less than 0.06%.
Trichococcus and Tetrasphaera genus were detected in all plants studied. Both genera have been reported to cause foaming problem in WWTPs as Nostocoida limicola morphotype and related to low temperatures [22, 23]. However, only Tetrasphaera in WWTP 2 and Trichococcus in WWTP 4 had relative abundances >3% which were considered as unusual percentages of total biomass, and therefore caused bulking and foaming processes [24].
Caldilinea was detected in the four plants, being the greater abundances in WWTP 2 (0.7-4.9%) and WWTP 4 (1.3-9%). This genus, known as Eikelboom's morphotype 0803, has been described as one that is able to grow both aerobically and anaerobically [25]. Caldilinea was found at high relative abundances from May to October in both plants coinciding with high temperature during the summer season. In WWTP 4, it increased again in December probably due to the spills from pear cannery campaign that was detected in November.
Thiothrix morphotype have been described as causative agents for sludge bulking [26]. Although this morphotype was found in all treatment plants, only WWTP 1 and 2 reached sufficient abundance to cause bulking problem (relative abundance of 6% in WWTP 1 and 4-6% in WWTP 2). Specifically, it was only during December that WWTP 1 showed a relative abundance of 6%, followed by less than 0.052% during the remaining period. In WWTP 2 however, the bulking lasted from May till October, which was associated with high sulfite (>2 mg/l), sulfate (>1,000 mg/l), chloride (>800 mg/l) and conductivity (>5,000 mg/l) as well as due to the presence of coloured dye (red and purple) that was detected at the plant inlet.
This study describes the changes of the microbial community by using metabarcoding analysis in 4 full-scale activated sludge WWTPs receiving discharges from different industries in Spain over one year period.
Proteobacteria phylum dominated most of the samples, but in some WWTPs the population shifted in bulking period from Proteobacteria dominated bacterial communities to Actinobacteria.
The phylum Firmicutes, which are versatile in degrading a big array of environmental substrates, dominated in WWTP 3 in August, associated with the start of peach canning campaign and with two spills detected in the plant entrance with high pH and conductivity.
In WWTP 2, which is characterized by textile discharges, Streptomyces species belonging to Actinomycetes capable of decolorize and mineralize textile dyes, were present at high relative abundance.
WWTP with IFAS system receiving cannery discharge showed the most diverse population of nitrifying bacteria, stating that attached biomass could facilitate the growth of nitrifying bacteria.
Nitrosomonas were the most abundant AOB in all WWTP, accounting up to 0.412% of the total sequences evaluated.
Nitrospira and Candidatus Nitrotoga coexisted in 3 of the 4 WWTPs analysed, with Nitrospira generally being the dominant NOB except in WWTP1 and WWTP2 where Candidatus Nitrotoga predominated over Nitrospira at different times of the year.
The decrease in nitrifying bacteria relative abundances suggested the loss of nitrogen removal efficiency. However, in some plants nearly zero relative abundance of the described species of AOB and NOB was detected without substantially affecting nitrogen removal efficiency. This suggests the presence of other undescribed genera of nitrifying bacteria as well as the possibility that NOBs can grow using the nitrite generated by nitrate-reducing bacteria.
Candidatus Microthrix was the major cause of sludge bulking in activated sludge plants with discharge from fruit handling, textile and leather industries, coinciding mainly with low temperatures but also with cleaning works of factories in the case of tanneries.
In WWTP receiving wastewater from canning industry, Caldilinea (0803 morphotype) was the main cause of sludge bulking, followed by Trichococcus (Nostocoida limicola morphotype), which was related to low temperatures.
In WWTP receiving textile discharges (WWTP 2), in addition to Ca. Microthrix, Thiothrix could be the source of bulking coinciding with high sulfite, sulfate, chloride and conductivity concentrations in plant inlet, together with Trichococcus and Caldilinea which had high abundance.
The data that support the findings of this study are available from the corresponding author, upon reasonable request.
We thank the funding from the European Commission through LIFE Programme (LIFE ENV16/ES/000390) and the WWTPs staff who have provided the data for this study.
The authors also thank to the owners of the WWTP where the samples were taken: Public Entity of Wastewater Sanitation (EPSAR) of the Valencian Community (Spain) attached to the Department of Agriculture, Rural Development, Climate and Ecological Transition and the Instituto Aragonés del Agua, attached to the Department of Agriculture, Livestock and Environment of the Government of Aragón (Spain).
[1] Wells, G.F., Park, H.D., Yeung, C.H., Eggleston, B., Francis, C.A., Criddle, C.S. (2009). Ammonia‐oxidizing communities in a highly aerated full‐scale activated sludge bioreactor: Betaproteobacterial dynamics and low relative abundance of Crenarchaea. Environmental Microbiology, 11(9): 2310-2328. http://dx.doi.org/10.1111/j.1462-2920.2009.01958.x
[2] Cydzik-Kwiatkowska, A., Zielińska, M. (2016). Bacterial communities in full-scale wastewater treatment systems. World Journal of Microbiology and Biotechnology, 32: 66. https://doi.org/10.1007/s11274-016-2012-9
[3] Zhang, B., Yu, Q., Yan, G., Zhu, H., Xu, X.Y., Zhu, L. (2018). Seasonal bacterial community succession in four typical wastewater treatment plants: Correlations between core microbes and process performance. Scientific Reports, 8(1): 4566. https://doi.org/10.1038/s41598-018-22683-1
[4] Xiao, Y.Y., De Araujo, C., Sze, C.C., Stuckey, D.C. (2015). Toxicity measurement in biological wastewater treatment processes: A review. Journal of Hazardous Materials, 286: 15-29. https://doi.org/10.1016/j.jhazmat.2014.12.033
[5] Islam, B.I., Musa, A.E., Ibrahim, E.H., Sharafa, S.A., Elfaki, B.M. (2014). Evaluation and characterization of tannery wastewater. Journal of Forest Products & Industries, 3(3): 141-150.
[6] Cortazar-Martínez, A., González-Ramírez, C.A., Coronel-Olivares, C., Escalante-Lozada, J.A., Castro-Rosas, J., Villagómez-Ibarra, J.R. (2012). Biotechnology applied to the degradation of textile industry dyes. Universidad y Ciencia, 28(2): 187-199.
[7] Klindworth, A., Pruesse, E., Schweer, T., Peplies, J., Quast, C., Horn, M., Glöckner, F.O. (2013). Evaluation of general 16S ribosomal RNA gene PCR primers for classical and next-generation sequencing-based diversity studies. Nucleic Acids Research, 41(1): e1. https://doi.org/10.1093/nar/gks808
[8] Altschul, S.F., Gish, W., Miller, W., Myers, E.W., Lipman, D.J. (1990). Basic local alignment search tool. Journal of Molecular Biology, 215(3): 403-410. https://doi.org/10.1016/S0022-2836(05)80360-2
[9] Wang, X.H., Hu, M., Xia, Y., Wen, X.H., Ding, K. (2012). Pyrosequencing analysis of bacterial diversity in 14 wastewater treatment systems in China. Applied and environmental microbiology, 78(19): 7042-7047. https://doi.org/10.1128/AEM.01617-12
[10] Hu, M., Wang, X.H., Wen, X.H., Xia, Y. (2012). Microbial community structures in different wastewater treatment plants as revealed by 454-pyrosequencing analysis. Bioresource Technology, 117: 72-79. https://doi.org/10.1016/j.biortech.2012.04.061
[11] Wang, J., Li, Q., Qi, R., Tandoi, V., Yang, M. (2014). Sludge bulking impact on relevant bacterial populations in a full-scale municipal wastewater treatment plant. Process Biochemistry, 49(12): 2258-2265. https://doi.org/10.1016/j.procbio.2014.08.005
[12] Liu, C., Li, H., Zhang, Y.Y., Si, D., Chen, Q.W. (2016). Evolution of microbial community along with increasing solid concentration during high-solids anaerobic digestion of sewage sludge. Bioresource Technology, 216: 87-94. https://doi.org/10.1016/j.biortech.2016.05.048
[13] Gonzalez-Martinez, A., Rodriguez-Sanchez, A., Lotti, T., Garcia-Ruiz, M.J., Osorio, F., Gonzalez-Lopez, J., Van Loosdrecht, M.C. (2016). Comparison of bacterial communities of conventional and A-stage activated sludge systems. Scientific Reports, 6(1): 18786. https://doi.org/10.1038/srep18786
[14] Park, H.D., Noguera, D.R. (2004). Evaluating the effect of dissolved oxygen on ammonia-oxidizing bacterial communities in activated sludge. Water Research, 38(14-15): 3275-3286. https://doi.org/10.1016/j.watres.2004.04.047
[15] Purkhold, U., Pommerening-Röser, A., Juretschko, S., Schmid, M.C., Koops, H.P., Wagner, M. (2000). Phylogeny of all recognized species of ammonia oxidizers based on comparative 16S rRNA and amoA sequence analysis: Implications for molecular diversity surveys. Applied and Environmental Microbiology, 66(12): 5368-5382. https://doi.org/10.1128/AEM.66.12.5368-5382.2000
[16] Wang, Z., Zhang, X.X., Lu, X., Liu, B., Li, Y., Long, C., Li, A. (2014). Abundance and diversity of bacterial nitrifiers and denitrifiers and their functional genes in tannery wastewater treatment plants revealed by high-throughput sequencing. PlOS ONE, 9(11): e113603. https://doi.org/10.1371/journal.pone.0113603
[17] Lücker, S., Schwarz, J., Gruber-Dorninger, C., Spieck, E., Wagner, M., Daims, H. (2015). Nitrotoga-like bacteria are previously unrecognized key nitrite oxidizers in full-scale wastewater treatment plants. The International Society for Microbial Ecology, 9(3): 708-720. https://doi.org/10.1038/ismej.2014.158
[18] Yao, Q., Peng, D.C. (2017). Nitrite oxidizing bacteria (NOB) dominating in nitrifying community in full-scale biological nutrient removal wastewater treatment plants. AMB Express, 7: 25. https://doi.org/10.1186/s13568-017-0328-y
[19] Baeza-Serrano, Á., Sempere, F., Oliver, N., Gutiérrez, P. and Fayos, G. (2022). Evolution of the activated sludge community of a wastewater treatment plant with industrial discharges. Ecology and the Environment, 259: 137-146. https://doi.org/10.2495/AWP220121
[20] Andreasen, K., Nielsen, P.H. (2000). Growth of Microthrix parvicella in nutrient removal activated sludge plants: Studies of in situ physiology. Water Research, 34(5): 1559-1569. https://doi.org/10.1016/S0043-1354(99)00319-X
[21] Knoop, S., Kunst, S. (1998). Influence of temperature and sludge loading on activated sludge settling, especially on Microthrix parvicella. Water Science and Technology, 37(4-5): 27-35. https://doi.org/10.1016/S0273-1223(98)00080-8
[22] Liu, J.R., Tanner, R.S., Schumann, P., Weiss, N., McKenzie, C.A., Janssen, P.H., Seviour, E.M., Lawson, P.A., Allen, T.D., Seviour, R.J. (2002). Emended description of the genus Trichococcus, description of Trichococcus collinsii sp. nov., and reclassification of Lactosphaera pasteurii as Trichococcus pasteurii comb. nov. and of Ruminococcus palustris as Trichococcus palustris comb. nov. in the low-G+C gram-positive bacteria. International Journal of Systematic and Evolutionary Microbiology, 52(4): 1113-1126. https://doi.org/10.1099/00207713-52-4-1113
[23] Seviour, R., Nielsen, P. H. (2010). Microbial ecology of activated sludge. The International Water Association Publishing, London, pp. 307.
[24] Daims, H., Lemmer, H., Nielsen, P. H. (2009). FISH handbook for biological wastewater treatment: Identification and quantification of microorganisms in activated sludge and biofilms by FISH. New York. The International Water Association Publishing, London. pp. 102458. http://library.oapen.org/handle/20.500.12657/25915.
[25] Kragelund, C., Thomsen, T.R., Mielczarek, A.T., Nielsen, P.H. (2011). Eikelboom's morphotype 0803 in activated sludge belongs to the genus Caldilinea in the phylum Chloroflexi. FEMS Microbiology Ecology, 76(3): 451-462. https://doi.org/10.1111/j.1574-6941.2011.01065.x
[26] Kanagawa, T., Kamagata, Y., Aruga, S., Kohno, T., Horn, M., Wagner, M. (2000). Phylogenetic analysis of and oligonucleotide probe development for Eikelboom type 021N filamentous bacteria isolated from bulking activated sludge. Applied and Environmental Microbiology, 66(11): 5043-5052. https://doi.org/10.1128/AEM.66.11.5043-5052.2000