Nafiseh Ghaffar Nia*![]() | Fariba Bahrami
| Fariba Bahrami![]() | Erkan Kaplanoglu
| Erkan Kaplanoglu![]() | Ahad Nasab
| Ahad Nasab![]()
© 2024 The authors. This article is published by IIETA and is licensed under the CC BY 4.0 license (http://creativecommons.org/licenses/by/4.0/).
OPEN ACCESS
Movement is one of the essential characteristics of living beings. Despite the diversity of animal species and the apparent differences, standard features exist between their movement systems that follow a particular pattern. The movements can mainly be divided into discrete and rhythmic categories controlled by the central nervous system. Scientists usually consider these two types of motion separately in the control system and use different methods and resources to produce and model them. Proposing a unified and comprehensive model for generating and controlling rhythmic and discrete movement with the same control system is more valuable, albeit challenging. This is essential because such a model would address a fundamental problem in the field of motor control, offering a holistic solution to understanding how living beings generate and control movement. A unified model could revolutionize various fields, including robotics, rehabilitation, and neuroscience, by providing a versatile framework applicable to various applications. In this study, we employed the Hodgkin-Huxley (HH) equations in our computational model; their suitability lies in their ability to capture the intricate dynamics of neural oscillations and the behavior of neural networks, making them an ideal choice for our investigation. Our comprehensive analysis of the model, factors influencing motion, and oscillation revealed crucial insights. We found that supraspinal input and motor neuron feedback, as the key motor control parameters, play pivotal roles in generating and controlling rhythmic and discrete movements. These findings contribute to our understanding of how the nervous system orchestrates both types of motion within a single framework. Developing a neuromechanical model capable of creating rhythmic and discrete movements holds promising implications. This research can potentially advance fields such as robotics, biomechanics, and rehabilitation by providing a unified framework for motor control. Moreover, understanding the switching mechanism between rhythmic and discrete states could lead to innovative strategies for designing more versatile and adaptive robotic systems and improving rehabilitation protocols for individuals with motor impairments.
rhythmic and discrete movements, CPG, neural oscillator, Hodgkin-Huxley equations, movement control, neuromechanical model
In neuroscience, the distinction between discrete and rhythmic movements is often studied in the context of motor control and motor learning [1]. Discrete movements are typically made up of a series of distinct, separate actions, such as reaching for a pen or writing on paper. In contrast, rhythmic movement is characterized by a repeated pattern or cycle of movement, such as the rhythm of running [2]. The interplay between rhythmic and discrete movements allows for the smooth, coordinated execution of complex actions, such as playing a musical instrument [3]. In the field of motor control, a discrete movement is a movement that is typically characterized by a distinct beginning and end, and it is often made up of a series of individual steps or sub-movements. On the other hand, rhythmic movement is characterized by repetitive motion patterns, often with a regular rhythm or beat.
The type of movement in every creature that can move is based on environmental conditions and body structures [4]. Various species' movement systems have standard features that follow a specific pattern [5]. The nervous system receives and processes multi-state motor neuron information and produces commands to control movement [6]. These commands are transmitted from the motor neurons in the spinal cord to the muscles and moving limbs, and through the motor neuron feedback, they return to the spinal cord for processing and completing actions [7]. Also, at the spinal cord level, some control operations are performed on the motor system, such as reflexes and adjustment of motor neuron weight to regulate the period, phase, and amplitude of the movement patterns [3].
Studying the neural basis of discrete and rhythmic movements can be helpful for various purposes, such as developing new technologies or therapies for people with movement disorders or improving human-machine interaction in robotics and other fields. They can be applied to stroke rehabilitation [8]; Therapists can design more effective rehabilitation programs and devices attached to patients and target specific neural pathways to help them regain motor function.
Both discrete and rhythmic movements are essential in the field of motor control, and they are used in various contexts. Our research aims to comprehensively understand how neural circuits, specifically central pattern generators (CPGs), control rhythmic and discrete movements and apply this knowledge to various practical applications. For instance, our findings hold great potential in developing therapies for individuals with movement disorders. By deciphering the neural mechanisms underlying discrete and rhythmic movements, we can design targeted interventions and rehabilitation strategies to improve these individuals' motor function and quality of life. In the realm of robotics, our research can revolutionize the design and control of robotic systems. The ability to generate both rhythmic and discrete movements using a unified neural model based on CPG can lead to more agile and adaptable robots capable of performing a wide range of tasks, from precise, discrete actions in manufacturing to dynamic, rhythmic motions in exploration and rescue missions. A CPG is a neural circuit that generates rhythmic patterns of movement and rhythmic activities, such as breathing, walking, and swimming, which do not require sensory input with timing information [9, 10, 11]. CPG is an oscillating network of different neurons found in animals’ spinal cords and brainstem [12]; it coordinates the activity of multiple muscles to produce the desired movement pattern. CPG's input is an elementary signal, but its output is complex, orderly, and coordinated. This point has received considerable attention in neuroscience for modeling motion [13]. The CPG-based model was also used as the controller of a humanoid robot to generate various walking patterns [14, 15]. Rhythmic movements generated by CPGs are often repetitive and stereotyped and can occur even without input from the brain. CPGs are found in many animals, including invertebrates and vertebrates, and are thought to have evolved to produce coordinated movement patterns for survival and reproduction. Indeed, CPG is a nonlinear oscillator as its primary function. It can only produce rhythmic motion and oscillations and send the output to the limb through flexor and extensor paths [16]. In other words, CPG’s outputs are usually rhythmic and repetitive activities. In this study, we have extracted both rhythmic and discrete output from the CPG model inspired by the model proposed by Markin et al. [17]. In the proposed model, CPG is located at the spinal cord level; it is symmetrical, meaning that the muscles' structure, the flexor, and the extensor are similar but in the opposite phase. This symmetry leads to the same changes in the stationary vertical phases [18].
By understanding the neural basis of discrete and rhythmic movements using CPG, we can advance our knowledge of how the brain controls and adapts to various motor tasks. We used dynamic systems analysis to discuss how this model works, what factors play an essential role in the generation of movement, how it can produce rhythmic and discrete movements, the role of motor neuron feedback, and whether motor neuron feedback is involved in the production of oscillation or not. Furthermore, if the primary supraspinal input at the surface of the brainstem is lost, can movement occur by amplifying other model components? We added the roles of the basal ganglia and cerebellum in producing movement and their impact on the supraspinal input to control movement. Finally, we implemented the effect of different control parameters in a proposed model to extract discrete and rhythmic movements.
Several models and theories have been highly influential in neuroscience regarding modeling movements. Some of the most notable ones include the CPG-based model [19], motor control theory [20], Hodgkin-Huxley model [21], and optimal control theory [22]. CPGs are neural circuits that generate rhythmic patterns of movement, such as walking and swimming, without the need for sensory input with timing information [19]. They have been pivotal in understanding and modeling rhythmic movements. CPGs are influential because they offer a neural basis for generating repetitive motions and have been applied in fields like robotics for developing locomotion controllers. On the other hand, motor control theories [20] focus on how the nervous system controls movements, including both discrete and rhythmic actions. The equilibrium point hypothesis, for instance, proposes that motor control is achieved by shifting the equilibrium point of muscles [23]. These theories have provided fundamental insights into the neural mechanisms underlying voluntary movements. While initially developed to understand the behavior of neurons, the Hodgkin-Huxley model's principles have been applied to study muscle activation and contraction, which is critical in movement generation [24]. Its mathematical rigor and ability to capture complex biological phenomena influence movement modeling. Optimal control theory borrowed from engineering and mathematics has been applied to study motor control [22]. It focuses on optimizing movement trajectories and has contributed to understanding how the nervous system might plan and execute movements efficiently.
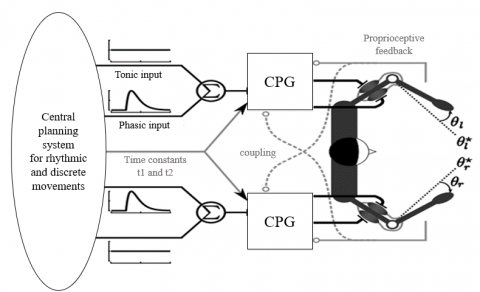
Furthermore, previous studies have proposed different views about motor neurons and rhythmic and discrete movements. One model claims that discrete movement is the fundamental and rhythmic movement is the concatenation of discrete movements [25, 26]. Another model states that rhythmic movement is the principal movement, and discrete movement is a truncated rhythmic movement [27, 28]. A third viewpoint mentions that rhythmic and discrete movements represent two different movement categories produced by the separate sections [29-31]. The third viewpoint is more acceptable based on research on the nervous system and motor behavior [32, 33]. In addition to these various theories and models, the motor control system requires precise modeling for further study. Also, there are different ways and structures for modeling the motor neuron system to produce different movements. Figure 1 demonstrates a model that combines electrical and mechanical systems to model rhythmic and discrete movements. This model uses a Matsuoka oscillator for the electrical part and a pendulum for the mechanical section [34].
At the beginning of the last century, scientists offered two interpretations for producing rhythmic movements. Charles Sherrington suggested one of these interpretations based on reflex loops [35, 36]. Sherrington believed that rhythmic movements result from the stimulation of reflex loops by motor neuron feedback, so motor neuron feedback ignites and switches between the different phases that make up a movement cycle. Another expression was proposed by Thomas Graham Brown, which was based on the centrality of rhythms [36]. Although this theory was challenged at its emergence, today, scientists believe that the CPG as biological neural circuits can produce rhythmic output in the absence of rhythmic input [37]. For example, stimulating a paralyzed fish (with spinal cord injuries) electrically or chemically will result in movement patterns called artificial movements, which are very similar to healthy movements [32]. Mock movements have also been observed in salamanders and adult frogs [38].
CPG has been seen in several animals, and research has shown that CPGs are broad networks consisting of several paired oscillating centers that produce rhythm [39, 40]. Although there is no need for motor neuron feedback to produce rhythmic output, this feedback plays an essential role in shaping the rhythmic pattern [41, 42]. The role of motor neuron feedback is critical for the CPG and body movements to be in harmony with each other.
Figure 1. A model for generating rhythmic and discrete movement by the Matsuoka oscillator [34]
In many vertebrates, electrical stimulation induces an area of the brain called the mesencephalic locomotor region (MLR) responsible for movement behavior [36]. At lower stimulation levels, a lower frequency of movements can be produced; correspondingly, higher stimulation levels result in a higher frequency of activities [43]. These observations show that CPG can generate complex movements by receiving simple inputs and switching between multiple phases.
CPG models are used in various robots and biomedical modeling [44]. Zhang et al. [45] proposed a new approach for designing a new actuator based on the CPG. The activity of the soft actuator is similar to octopuses’ muscles with no solid skeletal support. The authors simulated the gentle actuator activities under different conditions and controlled movements by adjusting the parameters of the CPG.
Moreover, CPG was used in designing six-legged Octopus robots [43]. It was also used in various swimming and crawling robots, such as snake robots [46, 47]. Furthermore, the CPG has been modeled in different levels of abstraction, from partial biophysical models to high-level abstract models. Partial biophysical models are based on the Hodgkin-Huxley model [48]. This neural model calculates how ions move within neural channels, how they affect the neurons' voltages and currents, and how nerve oscillations occur inside small neural circuits. While a large body of research has focused on the activity of single neurons, another group has focused on the dynamic properties of larger neural circuits. Also, rhythmic models use simple modeling of neurons, including the leaky receptor integral [49]. These models focus on how oscillating activity causes the properties of the neural network and how inter-neural connections synchronize different neural circuits.
Notably, there is an ongoing debate about whether motor control is hierarchical, with higher brain areas controlling lower-level movements, or if it involves distributed control across multiple levels simultaneously [50]. Understanding the nature of this control hierarchy remains a topic of discussion. Another debate revolves around the relative importance of feedback (sensory information during movement) versus feedforward (preplanned motor commands) control in movement generation and adjustment. This has implications for how we understand motor learning and adaptation. Also, the neural mechanisms underlying complex, coordinated movements, such as playing a musical instrument or skilled sports performance, are not fully understood [51]. There are debates about how these movements rely on specialized brain regions or result from extensive research and learning [52].
This research aims to contribute to these discussions by proposing a unified neuromechanical control model for generating and controlling both rhythmic and discrete movements. We provide insights into motor control's hierarchical or distributed nature by empirically demonstrating how a single model can account for diverse movement types. Additionally, our exploration of the role of supraspinal input, motor neuron feedback, and the impact of motor control parameters on movement generation can shed light on the debate regarding feedback versus feedforward control. Our study's findings can help delineate the neural mechanisms of switching between discrete and rhythmic states.
This model is designed with biological plausibility in mind. It includes various components, such as rhythm-generator neurons, motor neurons, interneurons, and muscle contractions, closely resembling the neural and mechanical elements found in living organisms. This biological relevance allows for insights into how real neural and muscular systems might generate and control movement. The model encompasses multiple neural and mechanical component layers, comprehensively representing the motor control system. By including rhythm generators, pattern-formation neurons, motor neurons, and feedback mechanisms, it accounts for various aspects of motor control. It can potentially provide a more holistic understanding of movement generation. The model's complexity and the inclusion of interneurons and inhibitory components make it suitable for investigating complex behaviors and transitions between discrete and rhythmic movements. Such flexibility can be crucial in addressing the research questions posed in the study.
The model's components, represented as neurons and muscles, can be described using mathematical equations, potentially allowing for analytical insights into the system's dynamics. Mathematical models often provide a precise framework for exploring how different parameters influence movement.
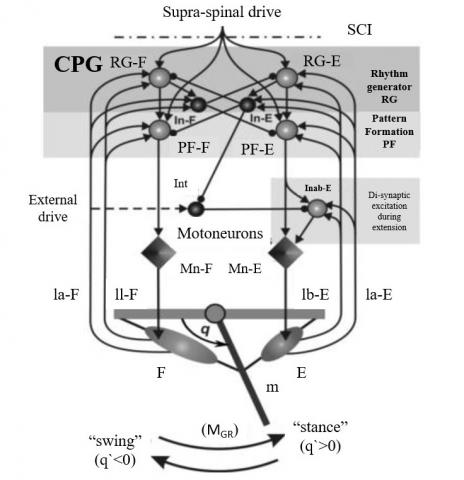
The primary model in this study is a simple neuro-mechanical model consisting of several layers, including rhythm-generator neurons (RG-F and RG-E), pattern-formation neurons (PF-F and PF-E), motor neurons (Mn-F and Mn-E), interneurons (In-F and In-E), inhibitory interneurons (Inab-E and Int), feedback neurons (Ia-F, IaE, II-F, and Ib-E) and two flexor and extensor contraction muscles that attach to the limb (F and E) (Figure 2).
Figure 2. Neuromechanical model and the relationship between its components [53]
Each neuron in this model represents a population of nerve cells that show the voltage difference between its membranes, represented by V, and the output of each neural population in the CPG layers denoted by f(v), which follows the nonlinear transformation below, where ${{v}_{\frac{1}{2}}}$ =−30 mV, k=3 mV for motor neuron, k=8 mV for other neurons, and Vth=−50 mV [53].
$f\left( v \right)=\left\{ \begin{matrix}
{}^{1}/{}_{\left( 1+\exp \left( -\left( \frac{V-{{v}_{\frac{1}{2}}}}{k} \right) \right) \right)~~~~~~~~~~~V\ge Vth} \\
0~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~Otherwise~~~~~~~~~~~~~~~~~~~~~~~~~~~~ \\
\end{matrix} \right.$
The neuron activation depends on whether or not their membrane voltage is greater than the threshold voltage (Vth). Two first-order differential equations below express dynamic relationships for rhythm-generator, pattern-formation, and motor neurons.
$C{{\dot{v}}_{i}}=-{{I}_{\text{Nap}}}\left( {{\nu }_{i}}{{h}_{i}} \right)-{{I}_{k}}\left( {{\nu }_{i}} \right)-{{I}_{\text{leak}}}\left( {{\nu }_{i}} \right)-{{I}_{\text{synE}}}\left( {{\nu }_{i}} \right)-{{I}_{\text{synI}}}\left( {{\nu }_{i}} \right)$.
${{\dot{h}}_{i}}=\frac{{{h}_{\infty }}\left( {{v}_{i}} \right)-{{h}_{i}}}{{{\tau }_{h}}\left( {{v}_{i}} \right)}$.
The internal neurons (In-F, In-E, Int, and Inab-E) equation is described by:
$C{{\dot{v}}_{i}}\overset{}{\mathop{=-{{I}_{leak}}\left( {{v}_{i}} \right)-{{I}_{synE}}\left( {{v}_{i}} \right)-{{I}_{synI}}\left( {{v}_{i}} \right)}}\,$.
where, vi is the voltage difference between the membranes of the neuron i, C is the membrane capacitance, INap is constant sodium current, Ik is potassium current, Ileak is leakage current, IsynE is the excitation input of the synapses entering neuron i, IsynI is the inhibitory input of the synapses entering neuron i, which follow equations below:
${{I}_{synE}}={{\bar{g}}_{synE}}\left( {{v}_{i}}-{{E}_{synE}} \right)\left( \mathop{\sum }_{j}^{n}aj,if\left( {{v}_{j}} \right)+{{c}_{i}}d+\mathop{\sum }_{k}^{m}{{w}_{i,k}}f\left( bk \right) \right)$.
${{I}_{Nap}}\left( {{v}_{i}},{{h}_{i}} \right)={{\bar{g}}_{nap}}{{m}_{nap}}{{h}_{i}}\left( {{v}_{i}}-{{E}_{Na}} \right)$. ${{m}_{nap}}={{m}_{nap}}p\left( {{v}_{i}} \right)$.
${{I}_{k}}\left( {{v}_{i}} \right)={{\bar{g}}_{k}}m_{k}^{4}\left( {{v}_{i}}-{{E}_{k}} \right),{{m}_{k}}={{m}_{k}}\left( {{v}_{i}} \right)$.
${{I}_{leak}}\left( {{v}_{i}} \right)={{\bar{g}}_{leak}}\left( {{v}_{i}}-{{E}_{leak}} \right)$.
${{I}_{synI}}={{\bar{g}}_{synI}}\left( {{v}_{i}}-{{E}_{synI}} \right)\mathop{\sum }_{j}^{n}{{b}_{j,i}}f\left( {{v}_{j}} \right)$
where, d is the supraspinal drive input to CPG neurons, ci is a factor scaling the drive d to neuron i, aj,i is the weight of the excitatory input from neuron j to neuron i, bj,i is the weight of the inhibitory input from neuron j to i, f(bk) denotes the kth feedback signal and$\text{ }\!\!~\!\!\text{ }$wi,k is the weight of that feedback (kth) to neuron i. More details and other parameters are provided by Spardy et al. [53, 54].
The supraspinal input and motor neuron feedback from the muscles are two critical inputs for CPGs [55]. Supraspinal input is information from higher brain regions like the motor cortex or basal ganglia that can affect a central pattern generator (CPG) in the nervous system, enabling the initiation, control, or modulation of rhythmic movements [56].
Motor neuron feedback refers to sensory information from muscles and tendons that informs the nervous system about body position and movement, helping maintain balance and coordinate motion [57]. This feedback is vital for maintaining balance and coordinating movement. It can also help to initiate movement, as the brain can use sensory information to determine the starting position of a movement and plan the appropriate motor commands to execute the desired movement. Both supraspinal drive and sensory feedback are essential for proper functioning CPGs and coordinating rhythmic activity patterns [58].
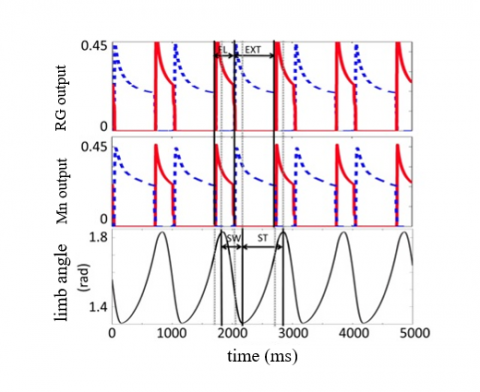
In addition, motor neuron feedback stimulates rhythm-producing and pattern-generating neurons, which result in increased oscillations. When rhythm-producing neurons are active in the path of the extensor muscles, rhythm-producing neurons in the direction of the flexor muscles are silent because, as mentioned before, this model is symmetrical. This fact also works for other neural layers. Figure 3 shows an example of rhythm-producing and motor neurons' activity. As shown in this diagram, when the neurons in the flexor muscle pathway (red line) are active, the neurons in the path of the extensor muscle are inactive (blue dashed line). This phase difference causes the extensor muscle to stop working while the flexor muscle starts working, and eventually, the limb begins to bend and contract. There are several models to create these symmetrical activities. For instance, a half-center oscillator CPG is commonly used to model the activation of opposing muscles. This is a specific type of CPG consisting of two interneurons that inhibit each other in a loop, creating a cyclical pattern of neural activity. It's often used to model opposing muscle activations. Each interneuron can alternately inhibit or excite the other, producing a cyclical pattern of neural activity that results in rhythmic motor output. The half-center oscillator CPGs can produce rhythmic muscle activation patterns without needing external stimuli or sensory feedback. They are also relatively simple to implement and can be used to investigate the basic principles of motor control.
Figure 3. The path of the flexor (FL) and extensor (EXT) muscles, which also work in the opposite phase [53]
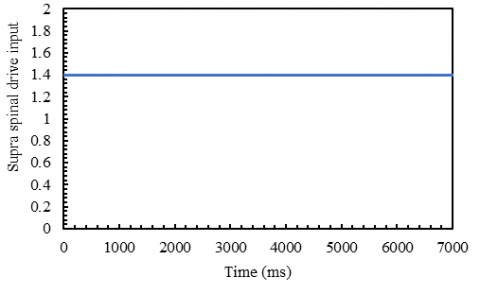
Figure 4. Supernatural input with a value of 1.4 (less than the stimulation threshold of 1.5)
Figure 5. Output (hand movement angle) for when we do not have proper supraspinal input in normal conditions
The CPG sends information to the limb muscles through the flexor and extensor pathways. The motor neuron feedback transmits information about the limb to the CPG, which indicates the angle of motion. It provides information to the nervous system to orient the body, regulate body movement, and communicate with the external environment. The supraspinal drive is a constant input from a higher level of the nervous system and primarily controls oscillation. The frequency fluctuations of CPG's output directly depend on this constant input strength.
In the following sections, we present the different parameters of this model that play an essential role in controlling and generating various movements applied to the neuromechanical system. Moreover, we examine the impact of multiple factors involved in the movement to indicate which factors could influence the fluctuation, period, and phase.
Our study shows that the system could oscillate with proper supraspinal input and adequate motor neuron feedback from the muscles. To be more exact, if the system fails to receive sufficient information from the supraspinal drive, it will not generate oscillation or action with the same conditions as before (Figures 4 and 5). Therefore, it is evident that the supraspinal input is essential in producing rhythmic information.
The remarkable point about the CPG is that simple signals are usually sufficient to stimulate and induct motion in the output [59]. Based on our research, the minimum amount of supraspinal input that can oscillate the system is 1.5, and higher values can lead to a higher frequency of movements. Likewise, low levels of stimulation reduce the frequency of movements (Figures 6 and 7).
Figure 6. The rhythmic movement's output angle when the supraspinal input amount is higher (4.7)
Figure 7. The output angle in the rhythmic movement when the amount of supraspinal input is small (2)
Without supraspinal input, the neurons cannot be activated; consequently, the limb has no motion. But could the motor neuron feedback be strong enough to move the motor limb? We removed the supraspinal input to answer this question and set the system with an initial amount of motor neuron feedback in the normal state. We observed that without input and a minimum amount of motor neuron feedback, the neuromechanical model could not produce oscillation, and the limb could not have movements. So, after increasing the motor neuron feedback coefficient, we observed that the intermediate neurons could be activated; this makes it possible to produce fluctuations in the output, and some fluctuations can be returned to the system. As mentioned earlier, when the input is zero, with the same initial conditions, the rhythm-generating neurons cannot be released from the dormant phase, and no activity can occur. It's important to note that the effects of feedback on a CPG can depend on the feedback's specific type and timing and the CPG's characteristics.
4.1 Examining the nullcline diagram of the model
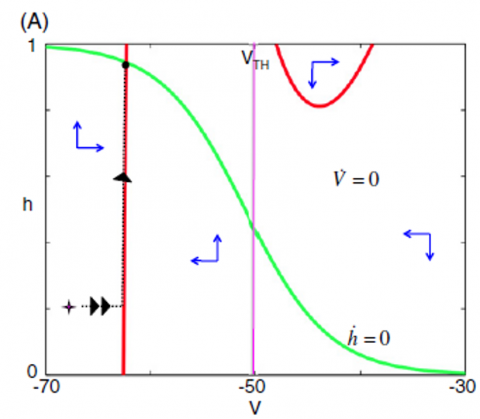
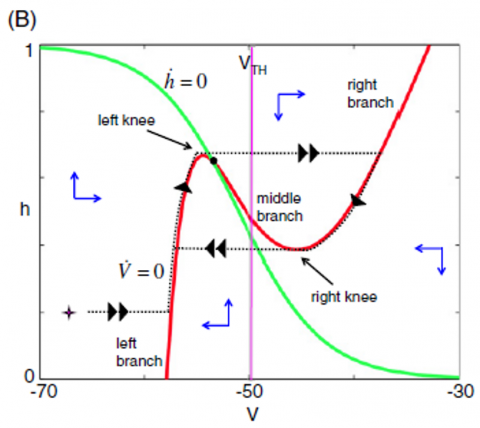
A nullcline diagram is a mathematical tool used to study the behavior of systems. In this context, it helps visualize how different inputs affect the activity of neurons. It is a map showing how neurons respond to various signals. The nullcline diagram in the phase space is an efficient mathematical concept used to analyze the dynamics of a system [60]. A system's phase space represents its state regarding its variables, such as position and velocity for a mechanical system or membrane potential and ionic concentrations for a biological system. The activity of neurons could be determined by their nullcline position in the phase space (V, h). In a biological system like a neuron, the nullcline position can be used to understand the equilibrium points and stability of the system. For example, when the nullcline position of the membrane potential intersects with that of a gating variable, it gives information about the system’s steady state. Additionally, the position of the nullcline can provide information about the system’s behavior, such as its stability and how it responds to perturbations.
Figure 8. An example of a system's nullclines graph for an input split. V-nullcline and h-nullcline are specified in pink and gray colors, respectively [53]
The intersection of the nullcline diagram, which is related to the system's input, shows different neuron states [61]. The neurons are inactive when the information is such that the h-nullcline and v-nullcline intersect in the left branch of the v-nullcline, Figure 8 (C). When h-nullclines and v-nullcline cross in the central unit of the v-nullcline, the neuron begins to produce bursts, Figure 8 (B); the subsystem enters a limit cycle and has burst as long as it is in this state. By changing the input, when the h-nullclines in the right branch of the v-nullcline intersect, the neuron begins to oscillate tonically, Figure 8 (A). Therefore, neurons can observe different activities by changing the input values; v-nullclines move to more positive voltages as we increase the feedback. Thus, activation in rhythm-generating neurons leads the intermediate neurons to exert a more robust input on the motor neurons, causing limb oscillations. When the information to the motor neurons is minimal, the oscillations disappear, and movement stops.
Rhythm-generating, pattern-generating, and motor neurons oscillate from their three input sources, including supraspinal input, interior neurons, and motor neuron feedback from the motor organ. If we remove one of these sources, the system will oscillate as long as the remaining two sources become more robust than in their normal state. So, rhythm-generating neurons could be activated with just two inputs, allowing intermediate neurons to exert more substantial input on motor neurons to produce oscillations in the limb. One of these two sources, the supraspinal information or motor neuron feedback, is always necessary to produce oscillations. However, these oscillations are not as strong as typical oscillations.
It is important to note that the nullcline diagram is just one aspect of the complex dynamics of a neuron, and other factors such as neurotransmitters, synaptic inputs, and intrinsic currents also play a role in determining the activity of a neuron.
Figure 9. Step input with a slight slope, 0.0001 in milliseconds
4.2 Examining different values for supraspinal drive input
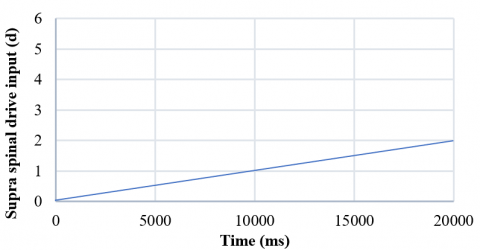
The supraspinal drive input is one of the most critical inputs to produce motion. According to this input, the neurons in CPG could be activated and run the motor control system, leading the system to oscillate. We observed constant fluctuations when we entered constant and stepped input to CPG as a supraspinal drive input. Also, when we applied the supraspinal input from the initial value of zero with a minimal step length (0.0001 ms), the system started oscillating as soon as the input's value (d) crossed 1.5. As time passes, these inputs and the output fluctuations also increase. Therefore, to change the output oscillation, we should change the amount of supraspinal drive input (Figures 9 and 10).
Figure 10. When the supra spinal drive input increases, the frequency of movement fluctuations also increases
4.3 Checking the role of motor neuron feedback in the absence of supraspinal drive input
When there is no supraspinal input, the system can only generate fluctuations in the output by increasing the motor neuron feedback strength. Still, as mentioned before, these oscillations are not as firm as they were in the normal state with supra spinal drive input (Figure 11).
Figure 11. Removing the supraspinal input and increasing the feedback power to generate oscillations
By examining the role of motor neuron feedback and its coefficient, we conclude that the most critical factor in motion is the motor neuron feedback coefficient. It can change the amplitude and frequency of movement and is an essential factor in controlling actions. The following section discusses the system dynamics scenarios through the bifurcation diagram.
4.4 Investigation of the effect of supraspinal input and the gain of motor neurons in the occurrence of bifurcation
Bifurcation is a sudden, significant change in a system's behavior. In this study, it refers to changes in neural activity patterns as inputs (supraspinal drive and motor neuron feedback) vary. Supraspinal input can influence the behavior of the nervous system and lead to changes in its activity. The increase in supraspinal input may lead to a bifurcation, which depends on a system’s limit cycle, input, and specific parameters, which causes a dramatic change in the behavior of a system. A limit cycle is a recurring pattern of behavior in a dynamic system. In this case, it represents the rhythmic movement of a limb controlled by the nervous system. This behavior is often seen in biological systems, such as the cardiac cycle, the neural activity that controls breathing, or rhythmic movements.
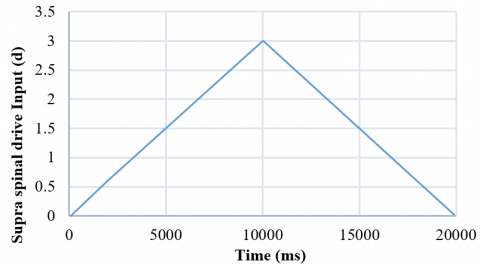
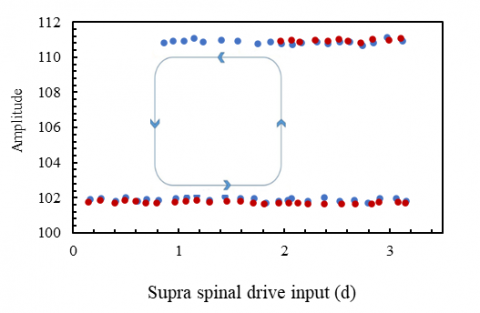
In the present study, the supraspinal input can lead the system to follow a bifurcation; when the input increases, the system enters the limit cycle at d=1.5. More severe fluctuations can be achieved by increasing the value of this input to 3. Then, by reducing this input with the same slight but negative slope (-0.003), the system at the point where d =1.0 goes out of the limit cycle and reaches a steady state with different values than when it started to increase. Therefore, the system has a limit cycle in which hysteresis has occurred. Fig. 13 shows the bifurcation diagram of the system when the supra-spinal drive input (d) follows Figure 12.
Figure 12. The supraspinal drive input is steadily increasing and decreasing to check the splitting in the system to evaluate the bifurcation diagram
Another essential factor in the system's bifurcation is increasing the motor neuron feedback coefficient, which leads to a significant decrease in the oscillation of the moving angle. Consequently, the bifurcation location will change, and the limit cycle will become smaller. As a result, when the amplitude of the oscillations decreases, the neurons enter and exit the limit cycle faster (Figures 13 and 14).
Figure 13. The system bifurcation diagram (MnE: 0.02)
When the supraspinal input in Figure 13 follows the diagram of Figure 12, the motor neuron feedback coefficient is 0.02, and other conditions are constant. The red and blue dots are the supraspinal input in the ascending and descending states
Figure 14. The system bifurcation diagram (MnE: 1)
When the supraspinal input in Figure 14 follows the diagram of Figure 12, the motor neuron feedback coefficient is 1, and other conditions are constant. The red and blue dots are the supraspinal input in the ascending and descending states.
By increasing the value of the motor neuron feedback coefficient, neurons exit the limit cycle more quickly. So, in this study, by examining the bifurcation diagram, we could understand that the supraspinal input and the motor neuron feedback coefficient can be the control parameters to observe the desired dynamic behavior in the system.
We should note that bifurcation diagrams are a tool for understanding the qualitative behavior of a system and may not capture all the details of the biological system, which is highly complex and multi-factorial.
Linear controller methods are based on the basic assumption of small-scale performance. This kind of controller cannot compensate for nonlinear effects in the system. On the other hand, nonlinear controllers cannot be easily implemented in the natural system due to the complexity of their structure. The choice of a proportional linear controller aligns with the study's goal of simplicity, given that all system parameters are known. This simplification allows for a clear understanding of the control system's behavior. Therefore, we can control the system by changing the proportional linear controller structure.
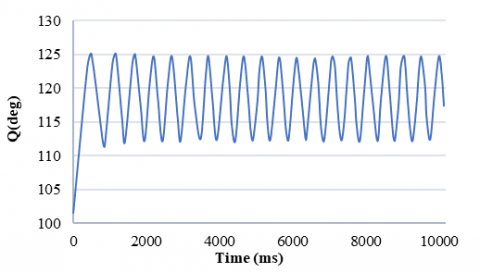
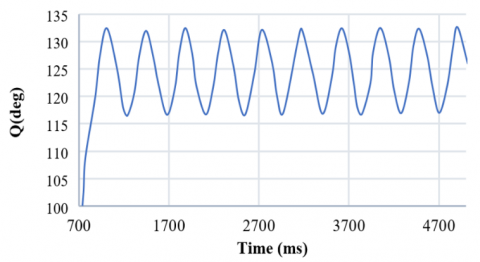
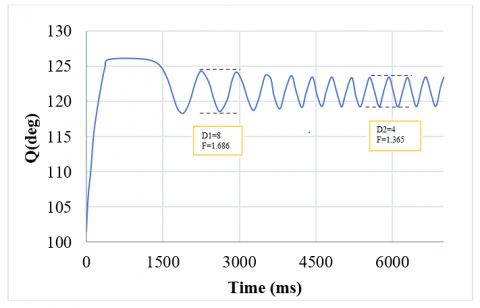
Since the physiological design of the nervous system can usually be influenced by strengthening or weakening its inputs, in this controller, there is a direct relationship between its inputs and outputs. In this system, the amplitude and the frequency of oscillations are limited. Therefore, we determined the maximum and minimum values of the movement's range and used the control system parameters, supraspinal input, and motor neuron feedback coefficient to design a controller. So, according to the effect of these two parameters, we achieved an appropriate control model derived from a proportional linear controller. However, there is always a persistent error between the desired value and the actual output in the zero-order system in a proportional controller. This error can be significantly reduced by increasing the controller gain, but it causes a delay, and the system returns to a steady state afterward. Therefore, we applied the exponential function to solve this problem. By using an exponential function and a training algorithm, the controller addresses the persistent error issue of linear controllers, which is crucial for achieving accurate and responsive control of limb movement. The controller's ability to limit the amplitude and frequency of oscillations reflects the natural constraints of limb movements, making it biologically plausible. As a result, this controller leads the system to respond quickly, reduces its overload, and increases the result accuracy to 99%. The system achieves the desired output in this simple model with very little error (less than 0.001). The resulting rhythmic motion is demonstrated in Figure 15.
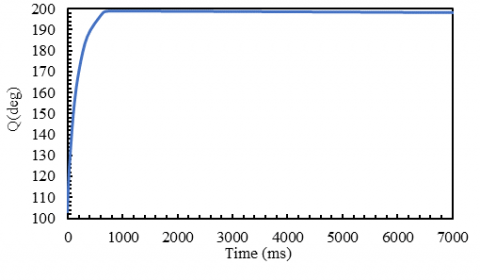
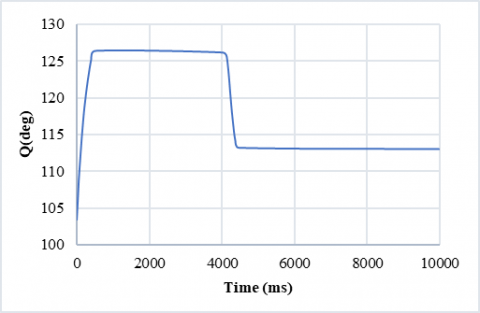
Discrete movements can also be modeled with this controller to simulate a limb’s motion from a certain angle to another desired angle. In this case, the frequency of activity is zero, and the control model repeats its algorithm to reach the desired output. Figure 16 shows an example of a discrete movement.
The chosen controller strikes a balance between simplicity and effectiveness. It leverages knowledge of the system's parameters and physiological principles to achieve accurate and responsive control of limb movements, making it suitable for the research's objectives.
Figure 15. The amplitude and frequency of the control model system decrease from 8 and 1.686 to 4 and 1.365, respectively
Figure 16. An example of discrete motion: the limb goes from 132 degrees to 115 degrees
Natural patterns always inspire researchers to design different systems. The movement of living organisms is a behavioral pattern considered in various studies. The central nervous system controls and produces motion activities through inhibitory and excitatory connections attached to muscles. This study examined the motor control system parameters through dynamics analysis and the bifurcation diagram. To this end, we evaluated the role of various components in the production and control of movement through bifurcation diagrams and system nullclines. Thus, we found that only two parameters, supraspinal input, and motor neuron feedback gain, are directly involved in this purpose. Finally, we could extract rhythmic and discrete movement by applying our simple controller to CPG, a nonlinear oscillator that can only generate a rhythmic motion with its initial structure. We should note focusing on minimal parameters (supraspinal input and motor neuron feedback gain) simplifies control design while maintaining effectiveness. Also, enabling rhythmic and discrete movements with a single control system advances the potential for developing more versatile and adaptable robotic systems. This simple controller could be used in robotic rehabilitation devices. These devices could assist patients in regaining motor control, improving their mobility and overall quality of life. Furthermore, this study paves the way for developing versatile robotic systems. Robots equipped with the controller described here could perform various tasks requiring rhythmic and discrete movements. This includes activities in industries such as manufacturing, healthcare, and agriculture.
[1] Bevilacqua, F., Boyer, E.O., Françoise, J., Houix, O., Susini, P., Roby-Brami, A., Hanneton, S. (2016). Sensori-motor learning with movement sonification: Perspectives from recent interdisciplinary studies. Frontiers in Neuroscience, 10: 385. https://doi.org/10.3389/fnins.2016.00385
[2] Sternad, D. (2008). Towards a unified theory of rhythmic and discrete movements—Behavioral, modeling and imaging results. In Coordination: Neural, Behavioral and Social Dynamics, pp. 105-133. https://doi.org/10.1007/978-3-540-74479-5_6
[3] Hogan, N., Sternad, D. (2007). On rhythmic and discrete movements: reflections, definitions and implications for motor control. Experimental Brain Research, 181: 13-30. https://doi.org/10.1007/s00221-007-0899-y
[4] Grillner, S., El Manira, A. (2019). Current principles of motor control, with special reference to vertebrate locomotion. Physiological Reviews, 100(1): 271-320. https://doi.org/10.1152/physrev.00015.2019
[5] Heermann, S. (2017). Neuroanatomy of the oculomotor system. Klin Monbl Augenheilkd, 234(11): 1334-1343.
[6] Nordin, A.D., Rymer, W.Z., Biewener, A.A., Schwartz, A.B., Chen, D., Horak, F.B. (2017). Biomechanics and neural control of movement, 20 years later: what have we learned and what has changed? Journal of Neuroengineering and Rehabilitation, 14: 1-11. https://doi.org/10.1186/s12984-017-0298-y
[7] Papale, A.E., Hooks, B.M. (2018). Circuit changes in motor cortex during motor skill learning. Neuroscience, 368: 283-297. https://doi.org/10.1016/j.neuroscience.2017.09.010
[8] Braun Janzen, T., Koshimori, Y., Richard, N.M., Thaut, M.H. (2022). Rhythm and music-based interventions in motor rehabilitation: Current evidence and future perspectives. Frontiers in Human Neuroscience, 15: 843. https://doi.org/10.3389/fnhum.2021.789467
[9] Berni, J., Pulver, S.R., Griffith, L.C., Bate, M. (2012). Autonomous circuitry for substrate exploration in freely moving Drosophila larvae. Current Biology, 22(20): 1861-1870.
[10] Berni, J. (2015). Genetic dissection of a regionally differentiated network for exploratory behavior in Drosophila larvae. Current Biology, 25(10): 1319-1326.
[11] Sims, D.W., Humphries, N.E., Hu, N., Medan, V., Berni, J. (2019). Optimal searching behaviour generated intrinsically by the central pattern generator for locomotion. Elife, 8: e50316. https://doi.org/10.7554/eLife.50316
[12] Steuer, I., Guertin, P.A. (2019). Central pattern generators in the brainstem and spinal cord: An overview of basic principles, similarities and differences. Reviews in the Neurosciences, 30(2): 107-164. https://doi.org/10.1515/revneuro-2017-0102
[13] Guertin, P.A. (2009). The mammalian central pattern generator for locomotion. Brain Research Reviews, 62(1): 45-56. https://doi.org/10.1016/j.brainresrev.2009.08.002
[14] Ijspeert, A.J. (2008). Central pattern generators for locomotion control in animals and robots: A review. Neural Networks, 21(4): 642-653. https://doi.org/10.1016/j.neunet.2008.03.014
[15] Nassour, J., Hénaff, P., Benouezdou, F., Cheng, G. (2014). Multi-layered multi-pattern CPG for adaptive locomotion of humanoid robots. Biological Cybernetics, 108: 291-303. https://doi.org/10.1007/s00422-014-0592-8
[16] Minassian, K., Hofstoetter, U.S., Dzeladini, F., Guertin, P.A., Ijspeert, A. (2017). The human central pattern generator for locomotion: Does it exist and contribute to walking? The Neuroscientist, 23(6): 649-663. https://doi.org/10.1177/1073858417699790
[17] Markin, S.N., Klishko, A.N., Shevtsova, N.A., Lemay, M.A., Prilutsky, B.I., Rybak, I.A. (2010). Afferent control of locomotor CPG: Insights from a simple neuromechanical model. Annals of the New York Academy of Sciences, 1198(1): 21-34. https://doi.org/10.1111/j.1749-6632.2010.05435.x
[18] Spardy, L.E., Markin, S.N., Shevtsova, N.A., Prilutsky, B.I., Rybak, I.A., Rubin, J.E. (2011). A dynamical systems analysis of afferent control in a neuromechanical model of locomotion: I. Rhythm generation. Journal of Neural Engineering, 8(6): 065003. https://doi.org/10.1088/1741-2560/8/6/065003
[19] Schmidt, A., Feldotto, B., Gumpert, T., Seidel, D., Albu-Schäffer, A., Stratmann, P. (2021). Adapting highly-dynamic compliant movements to changing environments: A benchmark comparison of reflex-vs. CPG-based control strategies. Frontiers in Neurorobotics, 15: 762431. https://doi.org/10.3389/fnbot.2021.762431
[20] Blakemore, R.L., Vuilleumier, P. (2017). An emotional call to action: Integrating affective neuroscience in models of motor control. Emotion Review, 9(4): 299-309. https://doi.org/10.1177/1754073916670020
[21] Catterall, W.A., Raman, I.M., Robinson, H.P., Sejnowski, T.J., Paulsen, O. (2012). The Hodgkin-Huxley heritage: From channels to circuits. Journal of Neuroscience, 32(41): 14064-14073. https://doi.org/10.1523/JNEUROSCI.3403-12.2012
[22] Wolpert, D.M., Ghahramani, Z. (2000). Computational principles of movement neuroscience. Nature Neuroscience, 3(11): 1212-1217. https://doi.org/10.1038/81497
[23] Latash, M. (2008). Evolution of motor control: From reflexes and motor programs to the equilibrium-point hypothesis. Journal of Human Kinetics, 19(2008): 3-24. https://doi.org/10.2478/v10078-008-0001-2
[24] Drukarch, B., Holland, H.A., Velichkov, M., Geurts, J.J., Voorn, P., Glas, G., de Regt, H.W. (2018). Thinking about the nerve impulse: A critical analysis of the electricity-centered conception of nerve excitability. Progress in Neurobiology, 169: 172-185. https://doi.org/10.1016/j.pneurobio.2018.06.009
[25] Cavallo, A., Bucchioni, G., Castiello, U., Becchio, C. (2013). Goal or movement? Action representation within the primary motor cortex. European Journal of Neuroscience, 38(10): 3507-3512. https://doi.org/10.1111/ejn.12343
[26] Latash, M.L., Levin, M.F., Scholz, J.P., Schöner, G. (2010). Motor control theories and their applications. Medicina, 46(6): 382. https://doi.org/10.3390/medicina46060054
[27] Jouaiti, M., Henaff, P. (2019). Real time movement classification in versatile CPG control. In Workshop on Robust Artificial Intelligence for Neurorobotics. https://hal.science/hal-02291647.
[28] Feldman, A.G. (1980). Superposition of motor programs—I. Rhythmic forearm movements in man. Neuroscience, 5(1): 81-90. https://doi.org/10.1016/0306-4522(80)90073-1
[29] Mottet, D., Bootsma, R. J. (1999). The dynamics of goal-directed rhythmical aiming. Biological Cybernetics, 80(4): 235-245. https://doi.org/10.1007/s004220050521
[30] Guiard, Y. (1993). On Fitts's and Hooke's laws: Simple harmonic movement in upper-limb cyclical aiming. Acta Psychologica, 82(1-3): 139-159. https://doi.org/10.1016/0001-6918(93)90009-G
[31] Guertin, P.A. (2009). The mammalian central pattern generator for locomotion. Brain Research Reviews, 62(1): 45-56. https://doi.org/10.1016/j.brainresrev.2009.08.002
[32] Brustein, E., Rossignol, S. (1999). Recovery of locomotion after ventral and ventrolateral spinal lesions in the cat. II. Effects of noradrenergic and serotoninergic drugs. Journal of Neurophysiology, 81(4): 1513-1530. https://doi.org/10.1152/jn.1999.81.4.1513
[33] Jouaiti, M., Henaff, P. (2018). CPG-based controllers can generate both discrete and rhythmic movements. In 2018 IEEE/RSJ International Conference on Intelligent Robots and Systems (IROS), Madrid, Spain, pp. 1212-1217. https://doi.org/10.1109/IROS.2018.8593889
[34] Ronsse, R., Sternad, D., Lefevre, P. (2009). A computational model for rhythmic and discrete movements in uni-and bimanual coordination. Neural Computation, 21(5): 1335-1370. https://doi.org/10.1162/neco.2008.03-08-720
[35] Burke, R.E. (2007). Sir Charles Sherrington's the integrative action of the nervous system: A centenary appreciation. Brain, 130(4): 887-894. https://doi.org/10.1093/brain/awm022
[36] Grillner, S., Ekeberg, Ö., El Manira, A., Lansner, A., Parker, D., Tegner, J., Wallen, P. (1998). Intrinsic function of a neuronal network—A vertebrate central pattern generator. Brain Research Reviews, 26(2-3): 184-197. https://doi.org/10.1016/S0165-0173(98)00002-2
[37] Pusuluri, K., Basodi, S., Shilnikov, A. (2020). Computational exposition of multistable rhythms in 4-cell neural circuits. Communications in Nonlinear Science and Numerical Simulation, 83: 105139. https://doi.org/10.1016/j.cnsns.2019.105139
[38] Ijspeert, A.J., Crespi, A., Ryczko, D., Cabelguen, J.M. (2007). From swimming to walking with a salamander robot driven by a spinal cord model. Science, 315(5817): 1416-1420. https://doi.org/10.1126/science.1138353
[39] Sims, D.W., Humphries, N.E., Hu, N., Medan, V., Berni, J. (2019). Optimal searching behaviour generated intrinsically by the central pattern generator for locomotion. Elife, 8: e50316. https://doi.org/10.7554/eLife.50316
[40] Steuer, I., Guertin, P.A. (2019). Central pattern generators in the brainstem and spinal cord: An overview of basic principles, similarities and differences. Reviews in the Neurosciences, 30(2): 107-164. https://doi.org/10.1515/revneuro-2017-0102
[41] Berkowitz, A. (2019). Expanding our horizons: Central pattern generation in the context of complex activity sequences. Journal of Experimental Biology, 222(20): jeb192054. https://doi.org/10.1242/jeb.192054
[42] Lodi, M., Shilnikov, A.L., Storace, M. (2019). Design principles for central pattern generators with preset rhythms. IEEE Transactions on Neural Networks and Learning Systems, 31(9): 3658-3669. https://doi.org/10.1109/TNNLS.2019.2945637
[43] Ijspeert, A.J., Crespi, A., Cabelguen, J.M. (2005). Simulation and robotics studies of salamander locomotion: applying neurobiological principles to the control of locomotion in robots. Neuroinformatics, 3: 171-195. https://doi.org/10.1385/NI:3:3:171
[44] Horcholle-Bossavit, G., Quenet, B. (2019). Neural network model of an amphibian ventilatory central pattern generator. Journal of Computational Neuroscience, 46(3): 299-320. https://doi.org/10.1007/s10827-019-00718-4
[45] Zhang, C., He, B., Ding, A., Xu, S., Wang, Z., Zhou, Y. (2019). Motion simulation of ionic liquid gel soft actuators based on CPG control. Computational Intelligence and Neuroscience, 2019: 8256723. https://doi.org/10.1155/2019/8256723
[46] Wang, M., Yu, J., Tan, M., Zhang, J. (2012). Multimodal swimming control of a robotic fish with pectoral fins using a CPG network. Chinese Science Bulletin, 57: 1209-1216. https://doi.org/10.1007/s11434-012-5005-6
[47] Wu, X., Ma, S. (2010). CPG-based control of serpentine locomotion of a snake-like robot. Mechatronics, 20(2): 326-334. https://doi.org/10.1016/j.mechatronics.2010.01.006
[48] Parker, J., Khwaja, R., Cymbalyuk, G. (2019). Asymmetric control of coexisting slow and fast rhythms in a multifunctional central pattern generator: A model study. Neurophysiology, 51: 390-399. https://doi.org/10.1007/s11062-020-09834-9
[49] Hellgren, J., Grillner, S., Lansner, A. (1992). Computer simulation of the segmental neural network generating locomotion in lamprey by using populations of network interneurons. Biological Cybernetics, 68(1): 1-13. https://doi.org/10.1007/BF00203132
[50] Grafton, S.T., Hamilton, A.F.D.C. (2007). Evidence for a distributed hierarchy of action representation in the brain. Human Movement Science, 26(4): 590-616. https://doi.org/10.1016/j.humov.2007.05.009
[51] Fairhurst, M.T., Janata, P., Keller, P.E. (2013). Being and feeling in sync with an adaptive virtual partner: Brain mechanisms underlying dynamic cooperativity. Cerebral Cortex, 23(11): 2592-2600. https://doi.org/10.1093/cercor/bhs243
[52] Lurie, D.J., Kessler, D., Bassett, D.S., Betzel, R.F., Breakspear, M., Kheilholz, S., Calhoun, V.D. (2020). Questions and controversies in the study of time-varying functional connectivity in resting fMRI. Network Neuroscience, 4(1): 30-69. https://doi.org/10.1162/netn_a_00116
[53] Spardy, L.E., Markin, S.N., Shevtsova, N.A., Prilutsky, B.I., Rybak, I.A., Rubin, J.E. (2011). A dynamical systems analysis of afferent control in a neuromechanical model of locomotion: I. Rhythm generation. Journal of Neural Engineering, 8(6): 065003. https://doi.org/10.1088/1741-2560/8/6/065003
[54] Spardy, L.E., Markin, S.N., Shevtsova, N.A., Prilutsky, B.I., Rybak, I.A., Rubin, J.E. (2011). A dynamical systems analysis of afferent control in a neuromechanical model of locomotion: II. Phase asymmetry. Journal of Neural Engineering, 8(6): 065004. https://doi.org/10.1088/1741-2560/8/6/065004
[55] Zehr, E.P. (2005). Neural control of rhythmic human movement: The common core hypothesis. Exercise and Sport Sciences Reviews, 33(1): 54-60.
[56] Turk, A.Z., Bishop, M., Adeck, A., SheikhBahaei, S. (2022). Astrocytic modulation of central pattern generating motor circuits. Glia, 70(8): 1506-1519. https://doi.org/10.1002/glia.24162
[57] Prochazka, A. (2015). Sensory control of normal movement and of movement aided by neural prostheses. Journal of Anatomy, 227(2): 167-177. https://doi.org/10.1111/joa.12311
[58] Grillner, S., El Manira, A. (2015). The intrinsic operation of the networks that make us locomote. Current Opinion in Neurobiology, 31: 244-249. https://doi.org/10.1016/j.conb.2015.01.003
[59] Ijspeert, A.J. (2008). Central pattern generators for locomotion control in animals and robots: A review. Neural Networks, 21(4): 642-653. https://doi.org/10.1016/j.neunet.2008.03.014
[60] Brauns, F., Halatek, J., Frey, E. (2020). Phase-space geometry of mass-conserving reaction-diffusion dynamics. Physical Review X, 10(4): 041036. https://doi.org/10.1103/PhysRevX.10.041036
[61] Ehsani, M., Jost, J. (2023). Scale free avalanches in excitatory-inhibitory populations of spiking neurons with conductance based synaptic currents. Journal of Computational Neuroscience, 51(1): 149-172. https://doi.org/10.1007/s10827-022-00838-4