Comparative Cultivation Performance, Biochemical Composition, and Molecular Identification of Three Sargassum Morphotypes from Lombok Waters, Indonesia

Nunik Cokrowati*![]() | Alis Mukhlis
| Alis Mukhlis![]() | Salnida Yuniarti Lumbessy
| Salnida Yuniarti Lumbessy![]() | Muhammad Marzuki
| Muhammad Marzuki![]() | Laily Fitriani Mulyani
| Laily Fitriani Mulyani![]() | Yuni Isniwati
| Yuni Isniwati![]()
© 2026 The authors. This article is published by IIETA and is licensed under the CC BY 4.0 license (http://creativecommons.org/licenses/by/4.0/).
OPEN ACCESS
Seedlings (propagules) of Sargassum collected from different geographic waters may exhibit higher metabolic activity and, under optimal light, temperature, and water-current conditions, achieve greater growth and biomass production. The objective of this study is to analyse the types of Sargassum sp. seeds that can produce optimal growth and biomass. This research was conducted from April 1 to December 1, 2024, at three sites (Ekas, Meninting, and Gerupuk) in Lombok, West Nusa Tenggara. The results show that Sargassum Ekas exhibited an absolute growth of 14.98 g, a specific growth rate of 3.45% per day, an alginate content of 0.508%, and a carbohydrate content of 4.89%. Sargassum Meninting exhibited absolute growth of 14.58 g, a growth rate of 3.47% per day, alginate content of 0.493%, and carbohydrate content of 6.45%. Sargassum Gerupuk exhibited absolute growth of 5 g, a specific growth rate of 3.13% per day, an alginate content of 0.378%, and a carbohydrate content of 7.34%. The three Sargassum species have a DNA identity percentage of 99.58-100%. This study concluded that Sargassum Ekas grew optimally in terms of biomass, alginate content, and biochemical composition. This is because seedlings from waters with extremely fluctuating water quality have developed this as an environmental adaptation.
cultivation, carbohydrates, growth, seaweed, Sargassum sp.
Sargassum sp. are brown macroalgae (class Phaeophyceae) that inhabit intertidal and subtidal zones, and several species are edible. The chemical composition of Sargassum sp. varies among species. In general, Sargassum sp. has several chemical contents, including water (17.29%), ash (22.56%), protein (10.64%), fat (1.99%), and carbohydrates (47.52%) [1]. The main component of brown algae is carbohydrates, while other components include protein, fat, ash, and water. Brown algae of the Sargassum genus have a monosaccharide composition consisting of 31-72% fucose, 5-31% galactose, and 3-9% xylose [2].
Sargassum biomass is influenced by multiple environmental factors. Temperature affects the rate of photosynthesis and respiration, whereas pH influences the availability of carbon and nutrients. Dissolved oxygen (DO) is necessary for dark respiration. The availability of nitrate and phosphate promotes biomass accumulation, while a deficiency in either of these elements causes growth limitations. Salinity controls cell osmotic pressure, which can inhibit ion absorption and reduce biomass. Light intensity (lux) affects energy absorption for photosynthesis, which is related to Sargassum biomass productivity [3].
The diverse geography of Indonesia's waters is an advantage for finding Sargassum seeds that will eventually reach maximum growth and biomass, an indication of long-term cultivation success. This study is based on the premise that Sargassum seeds originating from waters with optimal light, temperature, and currents have more active metabolism, resulting in higher growth rates and biomass production. The quality of these seeds also reduces physiological stress, leading to better seed viability and more stable growth rates throughout the cultivation period. In addition, the results of this study can be used as a basis for determining selection strategies and the movement of seed sources between regions, so that seaweed cultivation does not rely solely on local seeds that may be less than optimal, but instead selects seeds from waters with environmental conditions that produce the best growth and biomass.
The waters of the Ekas in southeast Lombok are a semi-enclosed bay connected to the open sea, with a temperature of around 28-30 ℃, salinity of 34-36 ppt, DO generally ≥ 5 mg/L, high transparency, and substrate currents dominated by sand and coral fragments, with little mud. Gerupuk Bay on the southern coast of Lombok is a sheltered bay with its mouth facing the Indian Ocean, currents of approximately 0.01–0.40 m/s, and substrates ranging from muddy sand to coral sand, with seagrass beds in shallow zones [2]. The waters of Meninting on the west coast of Lombok are influenced by the Meninting River estuary near the mouth, with low salinity, high turbidity, and a substrate of mud and muddy sand, transitioning to sand/coral sand. These different locations necessitate research and analysis of different seed sources originating from locations with different water temperatures (℃), pH, DO (ppm), ammonia (mg/L), nitrate (mg/L), phosphate (mg/L), salinity (ppt), and light intensity (lux) that produce seeds with optimal absolute growth, specific growth rate, alginate content, and carbohydrate content.
2.1 Study area and period
This study was conducted from April 1, 2024, to August 21, 2025, in Gerupuk Bay, Central Lombok, West Nusa Tenggara (Figure 1).

Figure 1. Research location in Gerupuk Bay, East Lombok, West Nusa Tenggara
2.2 Tools and materials
The supporting tools and equipment used are cultivation equipment and water quality measurement equipment. These tools include snorkeling goggles, manual scales, sample-measuring boards, Dissolved Oxygen meters, refractometers, Secchi disks, pH pens, boats, microscopes, cameras, hand tally counters, cool boxes, lux meters, scissors, knives, and reef boots.
The materials needed are Sargassum sp. seeds with an initial weight of 50 grams, base stakes, five mm-diameter twine, onion sacks, 50 kg sacks, raffia rope, mica tags, nitrate kits, ammonia kits, phosphate kits, alginate analysis materials, proximate analysis materials, dropper pipettes, and microscope slides.
2.3 Research method
This study used an experimental method conducted in marine waters (field). This study used three seed source groups and three replicates. The seed source group in this study was different types of Sargassum sp. seeds. The number of ropes used was the number of replicates in this treatment. The parameters measured in this study were growth, carbohydrate content, alginate content, and water quality. Figure 2 shows the design of the treatment and repetition layout.
Three Sargassum seed sources in Lombok have different characteristics: Ekas (East Lombok), which is semi-open, a deep bay with white sand, dominated by Andisol and Alluvial soils with fine-to-medium sand texture, and powerful reef break waves. The seawater quality meets the quality standards (KLH No. 51/2004). It is suitable for lobster and seaweed cultivation, with a moderate water-retention capacity that depends on coastal vegetation to prevent erosion. This location has two main river estuaries (Awang River Estuary and Kelongkong River Estuary) that carry natural nutrients to the bay, especially during the rainy season, and is dominated by a landscape of steep pyroclastic cliffs. Meninting (West Lombok) is an open coast facing directly onto the Lombok Strait, with volcanic black sand, alluvial soil (river deposits), and seasonal river-mouth break-wave types. Its water parameters indicate a pH of 8.0-8.2 and DO > 6 mg/L, which are ideal for underwater tourism. It is often studied due to the high concentration of suspended sediment from the Meninting River, especially during rainfall, and to the downstream area's high flood vulnerability, driven by the land slope and the type of alluvial soil. Its position is downstream of the Meninting watershed with volcanic sediments, and is dominated by gentle coastal alluvial plains with wide river confluences. Gerupuk (Central Lombok) is a hilly bay with white sand, brown Mediterranean soil, and sandy loam Grumusol, especially in mangrove areas and multi-level reef break wave types. The seawater quality shows stable salinity and pH, making it a suitable natural habitat for marine aquaculture, with runoff from surrounding river basins carrying nitrate and phosphate. The Mediterranean soil in this location has good water-retention capacity, but it is prone to drought if land cover is reduced. It has no large river estuaries, and its landscape is dominated by coral hills and headlands that protect the coast from strong winds.

2.3.1 DNA extraction and amplification
DNA extraction of Sargassum sp. samples from different regions was performed using the Quick-DNA Magbead Plus Kit (Zymo Research, D4082). The DNA extraction protocol was adjusted to match the kit's instructions for the Sargassum sp. DNA was then subjected to DNA quantity and quality tests to ensure it was suitable for molecular identification, of good quality, and optimal for DNA amplification.
The DNA amplification process was carried out using the Polymerase Chain Reaction (PCR) method. The gene amplification process was conducted using the KOD FX Neo kit (Toyobo, KFX-201) and a pair of primers for ITS2 gene amplification with the primer sequence 5.8S BF 5'-CGA TGA AGA ACG CAG CGA AAT GCG AT-3' and 25BR2 5'-TCC TCC GCT TAG TAT ATG CTT AA-3' [4]. The PCR program was as follows: initial denaturation at 95 ℃ for 3 minutes, followed by 35 cycles of denaturation at 95 ℃ for 15 seconds, annealing at 60 ℃ for 30 seconds, extension at 72 °C for 45 seconds, and final extension at 72 ℃ for 5 minutes. The amplification results were visualized using 1.5% agarose gel electrophoresis. PT Genetika Sains Indonesia then sequenced the targeted amplicon at 1st BASE. The test was conducted only once, namely for each seedling originating from a different region.
2.3.2 Molecular data analysis
The sequencing data were aligned using Molecular Evolutionary Genetic Analysis (MEGA) software. The identity of the nucleotide base sequence from each sample was analyzed using the Basic Local Alignment Search Tool (BLAST) web server at the National Center for Biotechnology Information (NCBI) [5]. The phylogenetic tree reconstruction was performed using MEGA with the neighbor-joining (NJ) method. Genetic distance was estimated using Kimura's two-parameter method based on 1000 bootstraps.
2.3.3 Alginate analysis
Alginate extraction was carried out by soaking 2 grams of dried seaweed in 100 mL of 0.33% HCl solution for 10 minutes to acid-pretreat the material. The seaweed was dried and blended in stages, after which 100 mL of Na₂CO₃ solution was added to dissolve the alginate under alkaline conditions whilst stirring until it thickened, ensuring a homogeneous alginate extraction. The thickened mixture was then dissolved in 600 mL of hot water and filtered to separate the liquid extract from the solid residue. The alginate was subsequently precipitated as calcium alginate by adding 50 mL of a 10% CaCl₂ solution to the filtrate, along with 100 mL of a 5% HCl solution to adjust the pH and facilitate gel formation. Next, 100 mL of distilled water was added until alginate clumps formed, which were then collected and dried in the sun to obtain the alginate extract in dry form.
2.3.4 Growth of seaweed
Absolute growth
Absolute growth is calculated using the following formula
W = Wt – W0
where,
W = Absolute growth in weight (g)
Wt = Average weight of test seaweed at the end of the experiment (g)
W0 = Average weight of test seaweed at the beginning of the experiment (g)
Specific growth rate
The daily growth rate is obtained using the following formula:
${SGR} \, (\%)=\frac{ln W_t-ln W_0}{t} \times 100$
where,
SGR = Specific growth rate (%)
Wt = Weight at last measurement (g)
W0 = Weight at stocking (g)
t = Duration of maintenance (days)
3.1 Seed types used
The Sargassum seedlings used in this study were collected from coastal waters around Lombok Island, West Nusa Tenggara, Indonesia. Three seed sources were evaluated: treatment A (Ekas), treatment B (Meninting), and treatment C (Gerupuk).
3.2 Absolute weight
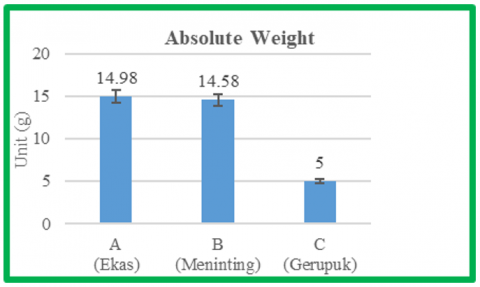
The cultivation results showed that the mean absolute growth differed among seed sources. Treatment A from Ekas waters was around 14.98 g, treatment B from Meninting waters was 14.58 g, and treatment C from Gerupuk waters was 5 g. The results of the absolute weight measurements are shown in Figure 3.
Figure 3. Absolute weight growth
The results of the ANOVA test at a significance level of 0.05 showed that different types of Sargassum sp. seaweed cultivated using the bottom stake method had a significant effect (P < 0.05) on the absolute weight growth of Sargassum. Based on further analysis (Duncan's test), treatment C was significantly different from treatments A and B, and treatment A was not significantly different from treatment B.
3.3 Specific growth rate
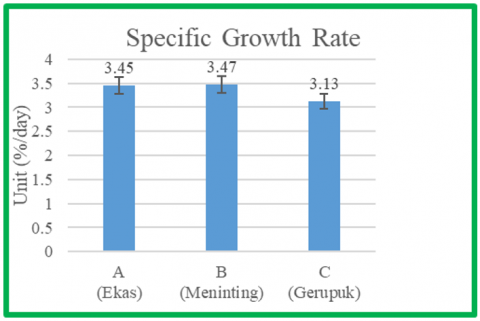
The results of this study indicate that the average specific growth rate of Sargassum cultivated for 17 days with different seedling types ranged from 3.13%/day to 3.47%/day. The highest average specific growth rate was found in treatment B at 3.47%/day, and the lowest specific growth rate was found in treatment C at 3.13%/day. The results of the growth rate measurements are shown in Figure 4.
Figure 4. Specific growth rate
3.4 Carbohydrate content
The chemical composition characteristics of the three species of wet brown algae Sargassum sp. that grow naturally in three waters of Lombok, West Nusa Tenggara (NTB) show varying values (Table 1). In general, brown algae (Sargassum) from the waters of Ekas Bay have the highest protein and ash content. Brown algae (Sargassum) from the Gerupuk seas have the highest fat and water content. Meanwhile, brown algae Sargassum from the waters of Meninting have the highest fiber and carbohydrate content. The chemical composition of seaweed can vary due to several environmental factors, including water temperature, salinity, light, nutrients, type, geographical location, and season [6].
Table 1. Chemical composition of wet brown algae Sargassum sp. in three waters of Lombok, West Nusa Tenggara (NTB)
|
Sampling Site |
Proximate Chemical Composition (%) (Wet Weight) |
|||||
|
Protein |
Fat |
Fiber |
Ash |
Water |
Carbohydrates |
|
|
Ekas |
2.36 |
0.29 |
0.54 |
7.65 |
84.81 |
4.89 |
|
Gerupuk |
1.39 |
0.34 |
0.48 |
2.08 |
89.26 |
6.45 |
|
Meninting |
2.03 |
0.17 |
0.86 |
2.69 |
86.91 |
7.34 |
3.5 ITS2 gene amplification and phylogenetic tree reconstruction
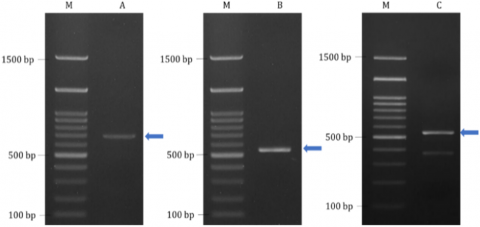
ITS2 gene amplification results for Sargassum sp. samples from the Ekas, Meninting, and Gerupuk show the presence of amplified bands in all samples (Figure 5). Meningting and Gerupuk show a clear single-band amplicon at ± 700 bp for Meningting and ± 550 bp for Gerupuk. Meanwhile, Sargassum samples from Ekas clearly showed two bands at ± 380 bp and ± 550 bp. The presence of bands in the electrophoresis results indicated successful amplification of the ITS2 gene in all samples, allowing further sequencing.
The sequencing process is necessary to identify the nucleotide sequence of the targeted gene and to determine the gene identity in the samples. These steps can reveal the genetic variations and characteristics of the observed samples. Based on the nucleotide base identity of threeMolecular identification based on the ITS2 gene shows that seedlings in groups A, B, and ℃ are genetically highly homologous to Sargassum polycystum, but there is a certain genetic distance (Figure 6), with group B (Meninting) being more distantly separated from the other two groups on the phylogenetic tree. As demonstrated by the similarity percentage between the Gerupuk area and S. polycystum, reaching 100%. Meanwhile, the Meninting has a similarity value of 99.85%, and the Ekas is 99.58% similar to S. polycystum.
Phylogenetic tree reconstruction of Sargassum sp. samples is essential for identifying relationships among samples and their taxonomic position across the three regions. The observed taxonomic position of Sargassum sp. is expected to facilitate further studies to assess its potential for optimal development and utilization.
The phylogenetic tree was reconstructed using MEGA with the neighbor-joining method. The phylogenetic results showed that Sargassum sp. from the Meninting were similar to S. yinggehaiense. Meanwhile, Gerupuk and Ekas formed a monophyletic branch that was similar to S. polycystum (Figure 6).
Table 2. ITS2 gene molecular identification of Sargassum sp. from three different sample locations
|
No. |
Sample |
Species Identification |
Query Cover (%) |
Percent Identity (%) |
Accession Number |
|
1 |
Meninting |
Sargassum polycystum |
71% |
99.85% |
JN038385.1 |
|
2 |
Gerupuk |
Sargassum polycystum |
100% |
100% |
KP096251.1 |
|
3 |
Ekas |
Sargassum polycystum |
100% |
99.58% |
MG731847.1 |

Figure 6. Phylogenetic tree reconstruction of Sargassum sp. samples from Meninting, Gerupuk, and Ekas compared with the GeneBank database using the Neighbor-joining (NJ) method
3.6 Alginate content
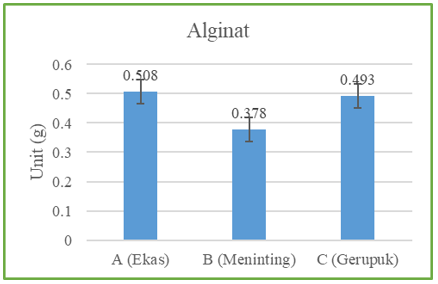
Figure 7 shows the range of alginate values obtained, namely Sargassum (A) 0.508%, Sargassum (C) 0.378%, and Sargassum (B) 0.493%. Based on the alginate analysis results, the lowest alginate content was found in Sargassum (C) and the highest in Sargassum (A).
Figure 7. Alginate content
3.7 Water quality
The values of the water quality parameters in Gerupuk Bay are as follows. Water quality parameters measured in Gerupuk Bay are presented in Table 3. The measurements were taken during Sargassum sp. cultivation using the bottom-stake method.
Table 3. Water quality parameters
|
Water Quality Parameters |
Range |
Day 17 |
|
Temperature (℃) |
32.2-32.8 |
32.2 |
|
pH |
8.2-8.27 |
8.2 |
|
Dissolved Oxygen (ppm) |
10.1-11.2 |
10.1 |
|
Ammonia (mg/L) |
1.5 |
1 |
|
Nitrate (mg/L) |
10 |
10 |
|
Phosphate (mg/L) |
1 |
1 |
|
Salinity (ppt) |
29 |
30 |
|
Light Intensity (lux) |
613 ´ 100 |
618 ´ 100 |
Sargassum seedlings from Ekas can demonstrate the best growth and highest alginate content because they originate from habitats with fluctuating environmental conditions (temperature, salinity, DO, light, currents) with sandy-rocky substrates that maximise nutrient absorption, so that environmental adaptation forms individuals with more efficient and productive physiology. Conversely, seedlings from Gerupuk can show the worst results because this bay has more heterogeneous conditions, including areas with muddy sand substrates and varying turbidity or currents, so that seedlings do not grow optimally and alginate synthesis is low, even though the research site is located here. This indicates that seedlings with good environmental adaptation can grow to their full potential compared to local seedlings originating from that environment. This is consistent with data on environmental differences in the seedling habitat that affect seedling physiology. Environmental adaptation leads to differences in growth capacity, as reflected in growth outcomes and alginate levels. This analysis is reinforced by environmental data from the seedling location, where the geographical base of Ekas waters makes it easier for seedlings to adapt and maximise their growth potential and alginate production.
4.1 Types of seedlings used
4.1.1 Sargassum (A)
This type is commonly found in the waters of Ekas Bay and West Nusa Tenggara. Figure 8 shows Sargassum (A) taken from the waters of Ekas Bay, East Lombok Regency.
This species has thalli shaped like thin leaves, with a paper-like texture and light weight. The thalli tend to cluster and form new thallus branches. This species has fruit-shaped thalli that function as air bubbles, allowing them to float in the water column [7]. This species can also be found in the waters of Sumbawa, Bima, and Dompu.
Figure 8. Sargassum (A)
4.1.2 Sargassum (B)
This species was obtained from the waters of Meninting Senggigi, West Lombok Regency. This species lives in the tidal zone on the coast and attaches itself to coral and rock substrates. In the third and fourth weeks of November and the first and second weeks of December, this species was abundant. Sargassum (B) is shown in Figure 9. This species has a thick, serrated leaf-shaped thallus. It also has a blade or fruit-shaped thallus that contains air.
Figure 9. Sargassum (B)
4.1.3 Sargassum (C)
Seedlings of this species were collected from the waters of Gerupuk Bay in Central Lombok Regency. This species has thin, elongated leaf-shaped thalli. Thallus growth tends to elongate in accordance with the height of the water column. Figure 10 shows Sargassum (C) cultivated in Gerupuk in this study.
Figure 10. Sargassum (C)
Sargassum (C) in the seedling phase or when still young, has many blades that resemble yellow bilateral flowers. In the adult phase, the talus is elongated and thin. This species can be found year-round in the waters of Gerupuk Bay.
4.2 Absolute weight
Based on the research conducted, the results show that the highest absolute weight was observed in treatment A, originating from Ekas waters, with an average absolute weight of 14.98 g, and the lowest in treatment C, originating from Gerupuk, with an average absolute weight of 5 g. Based on the results, the absolute weight gain in treatments A and B was classified as good. However, this was significantly different from treatment C. The difference in absolute weight between treatments was suspected to be due to their environmental adaptability. Treatments A and B had better adaptability and resistance to movement and water currents, resulting in more optimal nutrient absorption and optimal growth. The low absolute weight in treatment C is thought to be due to suboptimal adaptation, leading to stress and reduced nutrient absorption, as indicated by broken stems, fallen leaves, and fruit. This is in line with the statement [8]. The adaptation process in the talus causes the talus that cannot adapt well to new water conditions to change color to pale white, become brittle, and even break at the tips, reducing the talus's weight. Furthermore, research has shown that seaweeds that do not adapt well may experience stress due to changes in environmental conditions [8]. In addition, seeds taken from gerupuk are old seeds, which means that the seeds are of poor quality, slow to grow, and susceptible to disease. This is because Sargassum seedlings generally cannot be distinguished physically, as both the thallus and leaves look the same.
4.3 Specific growth rate
Based on the results obtained, the specific growth rate for each treatment was 3.14-3.47 g, which is considered good. This is reinforced by the statement [9] that daily seaweed growth above 2%/day indicates optimal growth, while growth below 2%/day indicates suboptimal growth. Water conditions are an essential factor in the growth of Sargassum sp, according to research [10]. The environment where Sargassum sp. algae grow is mainly in clear waters, with substrates such as coral reefs, dead coral, volcanic rocks, and massive objects on the bottom. Sargassum sp. algae grow in intertidal and subtidal areas, as well as in coastal areas with large waves and strong currents. The depth for growth is 0.5-10 m. The water conditions in which this research was conducted are consistent with those in which seaweed grows. This is supported by the study [11], which states that the waters in Gerupuk Bay are highly conducive to seaweed cultivation. In addition, the factors of time and place of cultivation significantly affect seaweed productivity. Furthermore, researchers [12, 13] argue that environmental pollution and nutrient availability alter the morphology and cytology of seaweeds, thereby affecting their growth and reproduction.
4.4 Carbohydrate content
Table 1 shows that the average wet weight content of Sargassum sp. brown algae growing naturally in three waters of Lombok ranges from 84.81 to 89.26%. The differences in moisture content among species of brown algae, Sargassum sp., that grow naturally in three waters of Lombok can be influenced by species, harvest age, and environmental conditions [14]. The moisture content of Sargassum sp. obtained in this study is in line with the moisture content of Sargassum sp. in the waters of Tapanuli, which ranges from 88.56 to 89.34% [15].
The protein content of the three brown algae species Sargassum sp. in this study ranged from 1.39 to 2.36% [16], states that brown seaweed contains 3-9% protein by wet weight, while red and green seaweed contain 6-20% protein by wet weight. In general, brown algae, including Sargassum, have a lower protein content than most brown algae. The low protein content of Sargassum is influenced by differences in species and season at the time of sampling [17].
The fat content of Sargassum sp. is not very high when compared to terrestrial plants. The average fat content of the three Sargassum sp. species in this study ranged from 0.17 to 0.34%. Marine macroalgae contain very little fat [16]. According to a study [18], the fat content of seaweed is generally less than 4% and is usually lower than that of terrestrial plants. The lowfat content is due to the storage of food reserves in plants as carbohydrates, especially polysaccharides, so that vegetable fats generally have a low percentage. Study [19] reported that macroalgae species living in tropical areas have a much lower fat content than subtropical species.
Ash content is the total mineral content contained in a material. The three Sargassum sp. species in this study had ash content values ranging from 2.08 to 7.65%. Ash (mineral) content is thought to be related to the absorption of mineral nutrients as an adaptation to marine environments with high mineral concentrations [15]. The absorption of seaweed minerals occurs throughout the entire surface of the thallus (not through the roots), resulting in effective uptake. The amount of minerals absorbed affects the ash content in seaweed tissue, resulting in high ash content [16]. The ash content of Sargassum sp. originating from Meninting waters is relatively high compared to other waters, presumably due to the high macro and micro mineral content of Meninting waters. This is supported by the physical and chemical factors of the Meninting waters, as oceanographic conditions remain suitable for Sargassum growth.
The carbohydrate content of the three brown algae species Sargassum sp. in this study ranged from 4.89 to 7.34%. Marine macroalgae and plants generally store their food reserves in the form of carbohydrates, especially polysaccharides. Polysaccharides are one of the components of carbohydrates. Carbohydrates are typically related to fiber in a material. High fiber content can be caused by high polysaccharide content in seaweed cells [14]. The results of this study show that the average fiber content of the three brown algae species Sargassum sp. ranges from 0.48 to 0.86%. The variation in crude fiber content is strongly influenced by photosynthesis, growth, and season. Meanwhile, seasonal changes can cause changes in environmental conditions that affect photosynthesis and nutrient absorption [20].
4.5 Alginate content
Sargassum is a type of brown seaweed found in warm waters worldwide, especially in tropical and subtropical regions. As a marine plant, Sargassum plays a role in the aquatic ecosystem and has potential for various applications, including as a source of alginate. According to research [21], Alginate is a compound extracted from the cell walls of brown algae, including Sargassum. Alginate has various uses, especially in the food, pharmaceutical, and other industries. Alginate has several benefits, including use as a thickener, emulsifier, and gel former in various food products such as ice cream, sauces, and other processed foods.
Alginate can also be used in the manufacture of tablets and capsules due to its ability to form gels. It can also be used to provide injection-based medicines. In addition, alginate can be used as a base material in the manufacture of tissue culture media for the growth of plants or animal cells [22]. Furthermore, alginate can be used in cosmetic products, such as face masks and scrubs, due to its ability to form an elastic film on the skin. Alginate can be used as a thickening agent or emulsifier in some beverages.
This study found the highest content in Sargassum (A), at 0.508 mg/L. Then, in Sargassum (B), the alginate content was 0.493 mg/L, and the lowest alginate content was found in Sargassum (C) with a value of 0.378 mg/L. This is supported by several factors, including differences in species, which may vary in their levels of alginate production. According to Kusumawati et al. [23], the dominant type of Sargassum in a region can affect the overall alginate content. In addition, water temperature, salinity, and nutritional conditions in the environment where Sargassum grows can affect alginate production [24]. Brown algae habitats that are more directly exposed to waves, live with roots firmly attached to coral, and have larger clumps have higher alginate levels.
Optimal environmental conditions can increase the production of this compound. Most types of algae, including Sargassum, have a specific growing season during which they can experience increased biomass production and the production of particular compounds. According to Roy [25], Alginate content may be higher during this growing season. Physiological factors, such as growth phase, plant age, and Sargassum health, can also influence alginate content. Healthy Sargassum in an active growth phase tends to have higher alginate content. According to Inem and Wasahua [26], the alginate content in brown seaweed can vary depending on the type, environmental conditions, the season when it is harvested, the extraction method used, and the part of the seaweed plant that is extracted.
Sunlight can also increase alginate content because light intensity affects photosynthesis. According to Hong et al. [27], sunlight exposure can affect photosynthesis, which in turn can impact the production of compounds such as alginate. Therefore, light intensity and duration (sunlight or photoperiod) can affect alginate content. In addition, the availability of nutrients, such as nitrogen and phosphorus, can play an essential role in the synthesis of secondary metabolites, including alginate. According to Zakaria et al. [28], seaweed that has adequate access to nutrients may tend to have higher alginate content.
Factors affecting low content in Sargassum include species and varieties of brown seaweed, which may differ in alginate content. According to Fenoradosoa et al. [29], the alginate content in Sargassum can vary between different species. Furthermore, the environmental conditions under which Sargassum is cultivated, or the Sargassum habitat, can affect alginate content. Water quality, such as temperature, DO, pH, nitrate, phosphate, and salinity at suboptimal cultivation sites, can affect Sargassum growth and cause stress, resulting in low alginate content and contributing to differences in specific compound content. This is reinforced by Samudra et al. [30], factors such as water temperature, nutrients, light intensity, and salinity can contribute to differences in the content of specific compounds.
In addition, the alginate content in Sargassum can vary throughout the year. According to Mas’Ud et al. [31], several factors, such as the duration of exposure to sunlight and the life cycle of marine plants, can affect the concentrations of specific compounds at different times of year. Furthermore, if Sargassum experiences poor growing conditions, such as exposure to pollution or an unsupportive environment, its alginate content may be low. External factors such as extreme weather, pollution, and climate change can also affect the growth and chemical composition of Sargassum.
4.6 Molecular character
Molecular-based identification is beneficial for clarifying an organism's taxonomic position. The presence of numerous morphological variations within a single alga can lead to errors during identification [32]. The numerous variations exhibited by an organism's morphological characteristics are understood to be influenced by environmental factors. Therefore, molecular identification is necessary to avoid errors.
The ITS2 gene, also known as a molecular barcode DNA region, was used in this study because it can identify organisms at the species level. The ITS2 gene was successfully amplified in several Sargassum spp. and has been reported to span 1250-1450 bp [33]. In the present study, the ITS2 gene amplicon was observed to have 380-700 base pairs. These results are aligned with the ITS2 gene amplification in Sargassum spp. samples from Yogyakarta [4].
Various factors, including DNA quality, primer design, inhibitor presence or absence, and PCR conditions, influence the success of ITS2 gene amplification by PCR. The absence of amplified DNA bands is suspected to be due to inhibitors present in the sample. Sargassum sp. is one species that is a source of alginate [34]. Alginate is a compound that inhibits the polymerase enzyme in the DNA amplification process, through the adsorption of Mg2+ from PCR reagents, trapping the polymerase enzyme and preventing it from catalyzing DNA strand elongation in the PCR process [35]. This necessitates a specialized method for isolating DNA from the Sargassum sp. species group to ensure that the alginate or other metabolite compounds produced within the individual do not interfere with molecular identification.
Sargassum sp. are a valuable source of bioenergy due to their high concentrations of organic compounds, minerals, carbohydrates, essential amino acids, and several inorganic compounds. Sargassum sp. also has a carbon and nitrogen ratio of 20:1 to 30:1 [36]. The high carbon-to-nitrogen ratio in Sargassum sp. is favorable for bioenergy production [37]. However, metabolite compounds in Sargassum sp. can vary by species. This makes molecular identification essential to ensure that the algae spreading in Indonesian waters are Sargassum sp., which have great potential for bioenergy production. This is reinforced by the many variations in the morphology of Sargassum sp., which often change in response to environmental factors, making the identification of Sargassum sp. difficult if done solely through morphological observation.
The molecular identification results for the ITS2 gene from the observed samples confirmed that Sargassum sp. from Gerupuk and Ekas were similar to S. polycystum, as indicated by BLAST and phylogenetic reconstruction. S. polycystum with Gerupuk and Ekas formed a monophyletic branch. When the same type of individual is in the same branch, or monophyletic, it can be assumed that they have a common ancestor [38]. These results differ from those of the Sargassum sp. sample from Meninting, which showed BLAST results similar to S. polycystum. Still, phylogenetic reconstruction showed that the sample was more closely related to S. yinggehaiense. The phylogenetic reconstruction reveals that Meninting forms a branch (clade) that is far apart from the Gerupuk and Ekas samples. This indicates there are genetic differences between Meninting and the two others.
The Sargassum sp. samples in the present study were morphologically identified as Sargassum aquifolium, Sargassum cristaefolium, and Sargassum hildebrantii. However, BLAST-based molecular identification confirmed that the observed Sargassum sp. was similar to S. polycystum. BLAST is an analytical tool for identifying the similarity of the nucleotide base sequence of an observed organism by comparing it with the gene database in NCBI. BLAST results indicate the identity of the nucleotide base sequence of the sample that was successfully sequenced [5]. Meanwhile, phylogenetic tree reconstruction indicates a relationship between observed samples with S. polycystum and S. yinggehaiense. This discrepancy between morphological and genetic identification results is often due to the genus Sargassum being known for high morphological plasticity. Therefore, identification of Sargassum sp. species based on morphological characteristics is considered less accurate [33]. Nevertheless, it remains necessary to employ morphological and molecular character-based approaches to identify species differences in Sargassum sp.
4.7 Water quality
The temperature obtained in the cultivation of Sargassum sp. using the bottom stake planting method in Gerupuk waters ranges from 32.2 ℃ to 332.8 ℃, which is the optimal temperature for the growth of Sargassum sp. According to Cokrowati et al. [39], the optimal water temperature for the growth of Sargassum sp. ranges from 26 ℃ to 30 ℃. Water temperatures that are too low or too high can cause poor growth for S. thunbergii. Enzymes that act as catalysts in the photosynthesis process cannot function properly if the water temperature is too low or too high.
The pH values obtained in this study ranged from 8.2 to 8.27. This is the optimal pH value for cultivating Sargassum sp. According to Hwang et al. [40], the appropriate pH for the growth of S. fulvellum ranges from 5 to 9. High pH conditions can cause osmotic stress in seaweed. This occurs when differences in ion concentrations inside and outside seaweed cells cause problems in water absorption.
The results of Dissolved Oxygen measurements in coastal waters showed values suitable for the growth of Sargassum sp. seaweed, ranging from 10.1 to 11.2 mg/L. The Dissolved Oxygen values in this study were optimal for cultivation. According to research [41], Dissolved Oxygen levels that support seaweed cultivation range from 8 to 11.77 mg/L.
The results of ammonia measurements in the waters around the planting site during the cultivation of Sargassum sp. ranged from 1 to 1.5 mg/L. These results indicate high ammonia levels in these waters. According to research [42], the optimal ammonia level for cultivation activities ranges from 0.04 to 0.06 mg/L.
The nitrate value obtained is 10 mg/L, a high concentration for the growth of sea grapes and seaweed. According to a study [43], nitrate levels in water ranging from 0.09 to 3.5 mg/L are good for seaweed growth in absorbing nutrients. In aquaculture, including Sargassum sp., nitrate (NO₃⁻) content is an essential nutrient needed for healthy growth and development.
The phosphate concentration obtained from Sargassum sp. cultivation is 1 mg/L, which is considered high. According to a study [42], the optimal phosphate value in water for seaweed cultivation is one µmol kg-1. Salinity is a measure of the amount of salt dissolved in seawater or other water solutions. The salinity values obtained in this study ranged from 29 to 30 ppt. This value is within the optimal salinity range for Sargassum cultivation. This is in line with the opinion of research [44] that Sargassum sp. seaweed can thrive in tropical waters with a salinity range of 29-33.5 mg/L.
The light intensity obtained in this study ranged from 613 × 100 Lux to 618 × 100 Lux. According to a study [43], the ideal light intensity for cultivation is around 5000 lux. Sufficient light intensity can increase seaweed growth rate. If the light intensity is insufficient, seaweed growth may slow down or even stop due to a lack of energy for photosynthesis [45].
Based on the results of research on the cultivation of Sargassum sp. seaweed using three types of seedlings from different waters, differences were found in growth, specific growth rate, genetic similarity, and alginate content. Treatment A, originating from Ekas waters, showed an average absolute weight growth of 14.98 g, followed by treatment B from Meninting waters at 14.58 g. In comparison, treatment C from Gerupuk waters produced the lowest increase at 5 g. The average specific growth rate ranged from 3.13 to 3.47% per day, with the highest value in treatment B at 3.47%/day and the lowest in treatment C at 3.13%/day. Genetic analysis showed that the Sargassum sample from Gerupuk had the highest similarity to Sargassum polycystum, namely 100%, while the sample from Meninting had a similarity of 99.85%, and that from Ekas had a similarity of 99.58%. In addition, the highest alginate content was found in Sargassum treatment A at 0.508%, followed by treatment B at 0.493%, and the lowest in treatment C at 0.378%. This study concludes that Sargassum from Ekas Bay (A) is the type that can grow optimally among the types studied.
Comparative cultivation performance in Ekas Bay shows high potential for local species such as Sargassum, which can be developed as a cultivation biostimulant and raw material for the food industry. Future research will focus on offshore cultivation techniques to reduce dependence on natural stocks, as well as the exploration of active compounds in alginate for global pharmaceutical applications.
The author would like to express gratitude to the University of Mataram for funding this research through its internal research funding for 2024.
[1] Sebastian, H.G.G. (2021). Growth and yield performance of rice cultivars (Oryza sativa L.) under seaweed extract and inorganic fertilizer. Plant Science Today, 8(4): 933-940. https://doi.org/10.14719/pst.2021.8.4.1072
[2] Cokrowati, N., Sunarwidhi, E., Raihanun, B., Astuti, N.K.P., et. al. (2024). Growth performance of Sargassum cristaefolium in Gerupuk Bay, Central Lombok. Media Akuatika, 9(1).
[3] Dolorosa, M.T., Nurjanah, Purwaningsih, S., Anwar, E. (2019). Utilization of Kappaphycus alvarezii and Sargassum plagyophyllum from Banten as cosmetic creams. IOP Conference Series: Earth and Environmental Science, 404(1): 012008. https://doi.org/10.1088/1755-1315/404/1/012008
[4] Saraswati, M.T., Adharini, R.I., Hardianto, E. (2024). The diversity of Sargassum spp. from the south coast of Yogyakarta, Indonesia, based on morphological characters and DNA barcoding ITS2 nrDNA. Biodiversitas, 25(8): 2733-2739. https://doi.org/10.13057/biodiv/d250846
[5] Madden, T. (2002). The BLAST sequence analysis tool. In National Center for Biotechnology Information (US).
[6] Herawati, D., Pudjiastuti, P. (2021). Effect of different solvents on the phytochemical compounds of Sargassum sp. from Yogyakarta and East Nusa Tenggara. Journal of Physics: Conference Series, 1783(1): 012001. https://doi.org/10.1088/1742-6596/1783/1/012001
[7] Cokrowati, N., Diniarti, N. (2019). Sargassum aquifolium components as growth-triging hormones for Eucheuma cottonii. Jurnal Biologi Tropis, 19(2): 316-321. https://doi.org/10.29303/jbt.v19i2.1107
[8] Maryunani, K., Lumbessy, S.Y., Cokrowati, N. (2025). Growth of Kappahycus alvarezii seaweed from tissue culture cultivated with different initial seed weights. Asian Journal of Fisheries and Aquatic Research, 27(2): 138-149. https://doi.org/10.9734/ajfar/2025/v27i2887
[9] Kusuma, N.P.D., Serihollo, L.G., Pratiwi, R., Amalo, P., Valentine, R.Y., Hariyadi, D.R., Usman, Z., Nomleni, A., Zulendra, M.F. (2025). Growth evaluation of the seaweed Kappaphycus striatus F. Schmitz (Solieriaceae: Rhodophyta) with mass selection method. Egyptian Journal of Aquatic Biology & Fisheries, 29(2). https://doi.org/10.21608/ejabf.2025.417084
[10] Lutfiawan, M., Karnan, Japa, L. (2015). Growth analysis of Sargassum sp. using different cultivation systems in Ekas Bay, East Lombok as enrichment material for the plant ecology course. Jurnal Biologi Tropis, 15(2): 135-144. https://eprints.unram.ac.id/9228/1/Muhammad%20Lutfiawan.pdf.
[11] Radiarta, N., Erlania, E. (2016). Performance of marine cultivation commodities in the integrated multi-trophic aquaculture (IMTA) system in Gerupuk Bay, Central Lombok, West Nusa Tenggara. Jurnal Riset Akuakultur, 11(1): 85-97. https://doi.org/10.15578/jra.11.1.2016.85-97
[12] Sulystyaningsih, N.D., Syamsuddin, R., Zainuddin, Z. (2019). The effect of depth and weight of Sargassum aquifolium on the level of ice ice attack and caragenan content in seaweed Kappaphycus alvarezii. Jurnal Riset Akuakultur, 14(1): 39-46. http://doi.org/10.15578/jra.14.1.2019.39-46
[13] Sahara, R., Herawati, V.E., Sudaryono, A. (2015). The effect of adding brown algae flour (Sargassum sp.) in feed on the growth and efficiency of feed utilization of catfish (Clarias sp.) fry. Journal of Aquaculture Management and Technology, 4(2): 1-8. https://ejournal3.undip.ac.id/index.php/jamt/article/view/8536.
[14] Adam, O.M., Nugraha, J., Machfoed, M.H., Turchan, A. (2021). In silico study on the promising active components of terpenoid and fucoidan from Sargassum sp. in inhibiting CGRP and TNF-α. Pharmacognosy Journal, 13(6): 1715-1719. https://doi.org/10.5530/pj.2021.13.221
[15] Dewinta, A.F., Susetya, I.E., Suriani, M. (2020). Nutritional profile of Sargassum sp. from Pane Island, Tapanuli Tengah as a component of functional food. Journal of Physics: Conference Series, 1542: 012040. https://doi.org/10.1088/1742-6596/1542/1/012040
[16] Handayani, T., Sutarno, Setyawan, A.D. (2004). Nutritional composition analysis of seaweed Sargassum crassifolium J. Agardh. Biofarmasi, 2(2): 45-52. https://doi.org/10.13057/biofar/f020201
[17] Fleurence, J. (1999). Seaweed proteins: Biochemical, nutritional aspects and potential uses. Trends in Food Science & Technology, 10(1): 25-28. https://doi.org/10.1016/S0924-2244(99)00015-1
[18] Kumar, A., Prakash, A., Dogra, S. (2011). Centella asiatica attenuates d-galactose-induced cognitive impairment, oxidative and mitochondrial dysfunction in mice. International Journal of Alzheimer’s Disease, 2011(1): 347569. https://doi.org/10.4061/2011/347569
[19] Garcia, J.S., Roldan, A., Palacios, V.M. (2016). Nutritional potential of four seaweed species collected in the Barbate Estuary (Gulf of Cadiz, Spain). Journal of Nutrition & Food Sciences, 6(3): 1000505 https://doi.org/10.4172/2155-9600.1000505
[20] Siddique, M.A.M., Bhuiyan, M.K.A., Khan, M.S.K. (2013). Nutritional composition and amino acid profile of a sub-tropical red seaweed Gelidium pusillum collected from St. Martin’s Island, Bangladesh. International Food Research Journal, 20(5): 2287-2292.
[21] Yanuhar, U., Suryanto, H., Amin, M., Binoj, J.S., Casuarina, I. (2024). Green synthesis of nano-copper oxide using Sargassum sp. functionalized in cellulose acetate membrane for dye adsorption. Global Journal of Environmental Science and Management, 10(4): 1933-1950. https://doi.org/10.22034/gjesm.2024.04.26
[22] Liu, J., Wu, S., Cheng, Y., Liu, Q., Su, L., Yang, Y., Zhang, X., Wu, M., Choi, J.I., Tong, H. (2021). Sargassum fusiforme alginate relieves hyperglycemia and modulates intestinal microbiota and metabolites in type 2 diabetic mice. Nutrients, 13(8): 2887. https://doi.org/10.3390/nu13082887
[23] Kusumawati, R., Basmal, J., Utomo, B.B. (2018). Physicochemical characteristics of sodium alginate extracted from Turbinaria sp. and Sargassum sp. Squalen Bulletin of Marine and Fisheries Postharvest and Biotechnology, 13(2): 79-84. http://doi.org/10.15578/squalen.v13i2.297.
[24] Hoang, T.C., Cole, A.J., Fotedar, R.K., O’Leary, M.J., Lomas, M.W., Roy, S. (2016). Seasonal changes in water quality and Sargassum biomass in southwest Australia. Marine Ecology Progress Series, 551: 63-79. https://doi.org/10.3354/meps11735
[25] Roy, S. (2020). Seaweed diversity within intertidal zone of Olaikuda and Vadakkadu, Rameshwaram, southeast coast of India. Heliyon, 6(7): e04585. https://doi.org/10.1016/j.heliyon.2020.e04585
[26] Inem, O., Wasahua, J. (2014). Potential types of brown algae in the coastal waters of Hutumuri Village, Ambon Island. Jurnal Ilmiah Agribisnis dan Perikanan (Agrikan UMMU-Ternate), 7(2): 39-45. https://doi.org/10.29239/j.agrikan.7.2.39-45
[27] Hong, M.H., Ma, Z.L., Wang, X.Y., Shen, Y.W., Mo, Z.Y., Wu, M.J., Chen, B.B., Zhang, T.T. (2021). Effects of light intensity and ammonium stress on photosynthesis in Sargassum fusiforme seedlings. Chemosphere, 273: 128605. https://doi.org/10.1016/j.chemosphere.2020.128605
[28] Zakaria, Z., Malik, A.A., Khairuddin, K., Ishak, M. (2023). The effect of fermentation duration on nutrition composition of seaweed (Sargassum sp.) liquid organic fertilizer. Journal of Aquaculture and Fish Health, 12(1): 1-11. https://doi.org/10.20473/jafh.v12i1.25669
[29] Fenoradosoa, T.A., Ali, G., Delattre, C., Laroche, C., Petit, E., Wadouachi, A., Michaud, P. (2010). Extraction and characterization of an alginate from the brown seaweed Sargassum turbinarioides Grunow. Journal of Applied Phycology, 22: 131-137. https://doi.org/10.1007/s10811-009-9432-y
[30] Samudra, A.G., Ramadhani, N., Pertiwi, R., Fitriani, D., Sanik, F., Burhan, A. (2023). Antihyperglycemic activity of nanoemulsion of brown algae (Sargassum sp.) ethanol extract in glucose tolerance test in male mice. Annales Pharmaceutiques Françaises, 81(3): 484-491. https://doi.org/10.1016/j.pharma.2022.11.011
[31] Mas’Ud, F., Santanumurti, M.B., Gabr, M.H., Jamal, M. T., Fanni, N. A., Saad, M. (2022). Lead (Pb) biosorption by intact biomass and alginate extract of Sargassum crassifolium originated from Gresik Regency waters. Ilmu Kelautan: Indonesian Journal of Marine Sciences, 27(2): 159-168. https://doi.org/10.14710/ik.ijms.27.2.159-168
[32] Widyartini, D.S., Widodo, P., Susanto, A.B. (2017). Thallus variation of Sargassum polycystum from Central Java, Indonesia. Biodiversitas, 18(3): 1004-1011. https://doi.org/10.13057/biodiv/d180319
[33] Yap-Dejeto, L.G., Fabillo, M., Sison-Mangus, M. (2022). Biodiversity of Sargassum (Fucales, Sargassaceae) from Eastern Samar (Philippines) inferred from nuclear ribosomal internal transcribed spacer (ITS) sequence data. Applied Phycology, 3(1): 422-434. https://doi.org/10.1080/26388081.2022.2119164
[34] Cokrowati, N., Prasedya, E.S., Ilhami, B.T.K., Hariadi, H., Jumat, M., Jayusri, Waang, D.C.S., Qoriasmadillah, W. (2022). Introduction of Sargassum sp. cultivation technology in Gerupuk, Central Lombok Regency. Jurnal Pengabdian Magister Pendidikan IPA, 5(4): 343-348. https://doi.org/10.29303/jpmpi.v5i4.2538
[35] Budiarto, B.R. (2018). The relationship between genotyping errors and the performance of nucleic acid amplification-based cancer molecular diagnostics. Biodidaktika: Jurnal Biologi dan Pembelajarannya, 13(2): 1-18. https://doi.org/10.30870/biodidaktika.v13i2.3279
[36] Murjani, J., Putri, R.M.S., Oktavia, Y., Ilhamdy, A.F. (2020). Biogas production from seaweeds Sargassum sp. Marinade, 3(1): 11-21. https://ojs.umrah.ac.id/index.php/marinade/article/view/2722/1116.
[37] Milledge, J.J., Harvey, P.J. (2016). Golden tides: Problem or golden opportunity? The valorisation of Sargassum from beach inundations. Journal of Marine Science and Engineering, 4(3): 60. https://doi.org/10.3390/jmse4030060
[38] Paterson, R.S., Rybczynski, N., Kohno, N., Maddin, H.C. (2020). A total evidence phylogenetic analysis of pinniped phylogeny and the possibility of parallel evolution within a monophyletic framework. Frontiers in Ecology and Evolution, 7: 457. https://doi.org/10.3389/fevo.2019.00457
[39] Cokrowati, N., Junaidi, M., Affandi, R.I., Sumsanto, M., Muahiddah, N., Anggraini, I.D., Marno, S., Asri, Y., Dwiyanti, S., Lumbessy, S.Y., Latifah, W., Fikri, R.A. (2024). The distribution, habitat characteristics, and bioenergy potential of Sargassum sp. in Indonesia. International Journal of Design & Nature and Ecodynamics, 19(6): 2049-2062. https://doi.org/10.18280/ijdne.190621
[40] Hwang, E.K., Yoo, H.C., Baek, J.M., Park, C.S. (2015). Effect of pH and salinity on the removal of phytal animals during summer cultivation of Sargassum fusiforme and Sargassum fulvellum in Korea. Journal of Applied Phycology, 27: 1985-1989. https://doi.org/10.1007/s10811-014-0511-3
[41] Li, H., Moon, H., Kang, E.J., Kim, M. et al. (2022). The diel and seasonal heterogeneity of carbonate chemistry and dissolved oxygen in three types of macroalgal habitats. Frontiers in Marine Science, 9. https://doi.org/10.3389/fmars.2022.857153
[42] Han, Y., Li, X., Li, X., Wang, S., Zhang, M., Li, M. (2025). Effects of acute and chronic ammonia exposure on survival, growth and intestinal microbiota composition of hybrid carp. Aquaculture Reports, 42: 102834. https://doi.org/10.1016/j.aqrep.2025.102834
[43] Novianti, D.N., Rejeki, S., Susilowati, T. (2015). The effect of different initial weights on the growth of Latoh Seaweed (Caulerpa lentillifera) Cultivated on the Bottom of a Jepara Pond. Journal of Aquaculture Management and Technology, 4(4): 67-73. https://www.neliti.com/publications/91996/pengaruh-bobot-awal-yang-berbeda-terhadap-pertumbuhan-rumput-laut-latoh-caulerpa.
[44] Ibrahim, A.M., Subiyanto, S., Ruswahyuni, R. (2014). The relationship between the density of Sargassum sp. seaweed and the abundance of Epifauna at Barakuda Beach, Kemojan Island, Karimunjawa Islands, Jepara. Jurnal Management of Aquatic Resources, 3(2): 36-44. https://www.neliti.com/id/publications/190669/hubungan-kerapatan-rumput-laut-sargassum-sp-dengan-kelimpahan-epifauna-di-pantai.
[45] Metsoviti, M.N., Papapolymerou, G., Karapanagiotidis, I.T., Katsoulas, N. (2019). Effect of light intensity and quality on growth rate and composition of Chlorella vulgaris. Plants, 9(1): 31. https://doi.org/10.3390/plants9010031