Hanan Shihab Ahmad*![]() | Saja Jamal Noman
| Saja Jamal Noman![]()
© 2024 The authors. This article is published by IIETA and is licensed under the CC BY 4.0 license (http://creativecommons.org/licenses/by/4.0/).
OPEN ACCESS
The present research explores the effects of Candida krusei yeast infection and chitosan treatment on rabbit liver and kidney functions. Utilizing male rabbits as a model, the research categorizes them into four distinct groups for clarity: the Candida krusei Infection Group, the Chitosan Treatment Group, the Combined C. krusei and Chitosan Group, and the Control Group. It was found that the C. krusei Infection Group and the Combined C. krusei and Chitosan Group had a significant increase in blood serum glucose levels as compared to the Control Group (P < 0.05). Comparatively, the Chitosan Treatment Group exhibited a significant reduction in glucose levels indicating a neutralizing impact on the hyperglycemia caused by C. krusei. The other finding is that the Group in the Chitosan Treatment 4 demonstrated significantly reduced levels of urea and uric acid in the blood serum than in the Group in the Control, which reflects a likely protection of kidney function (P < 0.05). In relation to protein metabolism, the Chitosan Treatment Group presented an elevation in total plasma protein levels and a reduction in serum albumin concentrations with significant differences (P < 0.05) when compared to the Control Group. The globulin levels analysis, in turn, confirmed these results, displaying a close tendency. Liver tissue sections histological examination reported that a hemorrhage within the parenchyma and hemolysis in the Chitosan Treatment Group, the C. krusei, and Nystatin Treated Groups showed inflammatory infiltrates, and extensive steatosis. The steatosis and lymphocytes infiltration were found in C. krusei Infection Group, in turn the focal lymphocytic infiltration and some necrotic cells were observed in Combined C. krusei and Chitosan Group. Kidney tissues in the Chitosan Treatment Group exhibited glomerular atrophy, while those of the C. krusei and Nystatin-Treated Groups showed lymphocyte infiltration and renal tubule dilation. The C. krusei Infection Group presented with renal tubule laceration and endothelial cell degeneration, whereas the Combined C. krusei and Chitosan Group had glomerular prolapse and early atrophy. All in all, in this study, the effects of C. krusei infection and chitosan therapy on glucose metabolisms, renal function and histological integrity of liver and kidney tissues in rabbits were revealed to be contrasting. The chitosan treatment provided resistance to the rabbits against the systemic effects of the C. krusei infection, showing its prophylactic role in both fungal infections and their systemic consequences. This type of knowledge reinforces the chitosan therapy possibility, and at the same time dictates the necessity of more clinical use and safety studies.
yeast infection by Candida krusei, chitosan treatment, functions of kidneys and liver, histological changes in liver and kidneys, levels of blood glucose, protein metabolism and drug resistance by fungus
In the fact, the liver and kidneys are the ones that are responsible for the metabolism and detoxification [1]. A few additional notes: In the liver, the cytochrome P450 system is one of the most important systems for the metabolism and removal of many toxins and chemicals [2], including drugs. Both the presence of P450-mediated interference and the loss of function of all members of the CYP superfamily may cause chemical sensitivity and difficulty in clearing the medications [3, 4]. Infections and toxins can destroy the liver's tissue, which usually neutralizes the impacts of metabolism or detoxification processes. It stresses on how good we take care of liver health [5]. The kidneys continuously filter the blood and eliminate waste that leads to urine. For kidney infection or toxins which can impair kidney tissue are contributors to creation of waste such as creatinine, urea, and others by this renal filtration system. In the last couple of decades, fungal diseases have become an epidemic, that's to say, more prominent and frightening. Along with conditions depressing immune function, general antibiotic abuse and others are the parts of the problem as well. One of the fields of medical research critical for the advancement of modern healthcare that should not be neglected is the development of safe and effective drugs against fungal infections. Protecting the life principles of vital organs particularly liver and kidneys through lifestyle factors (diet, exercise, stress management, etc.), trying to use unnecessary medications/toxics to the lowest would be an effective way to live a healthy life. In-teal metabolism, the function of all organs is undermined, which thereby leads to overall health and resistance of the body. Candida krusei is the major challenge in clinical setting as it exhibits excessive resistance to most of the antifungal drugs [6]. Such resistance requires the study of alternative or adjunctive therapies to the current antifungal drugs which often produce harmful side effects or are not effective against drug resistant strains [7]. Chitosan is a natural biopolymer derived from chitin, which has various therapeutic potential such as hypocholesterolemia effects and an ability to form chemical bonds with negatively charged molecules like fats and proteins [8]. This potential provides a precedent for the anti-fungal effects of chitosan on liver and renal functions by the alteration of the body’s metabolism and toxin elimination [9]. The objective of this study is to fill in the missing information virtually on the effect of chitosan therapy on the activity of liver and kidneys during the infections provoked by Candida krusei [10]. We speculate that this chitosan could possess hepatonephric protective properties from the pathogenic effects of C. krusei on these organs, which provide a new prospective of fungal infections and their consequences on metabolic health [11-14].
Research employed chitosan got from the Chinese firm (Xi'an), dissolving it in distilled liquid at a concentration of 150.0 mg/100.0 ml for its intended application. Given chitosan's solubility profile, an acidic solution (acetic acid 1%) was used to aid dissolution, adjusting the pH to 5.5, ensuring complete solubilization [15]. Candida krusei ATCC 6258 was utilized for this study, cultured on Sabouraud Dextrose Agar and maintained at 37℃ with a 24-hour incubation period to ensure optimal growth conditions.
Experiment Design: Twenty-five native male rabbits were systematically assigned into five groups, each consisting of five animals. The groups were as follows:
Collecting of Blood Samples: Following the dosing period, animals were anesthetized with chloroform for blood collection via cardiac puncture using a 5ml medical syringe. Blood samples were collected into clean, dry plastic tubes without anticoagulant to allow for serum separation. After standing for 15 minutes, the samples were centrifuged at 3000 rpm for 15 minutes. The serum was then withdrawn using a micropipette and stored at -20℃ until further analysis.
Yeast Suspension Preparation: The "yeast suspension" was prepared by inoculating a loopful of C. krusei culture into 5.0 ml of sterile saline solution, adjusting the concentration to 1.50 × 108 cells/ml, a typical range for infective dose studies [16]. Anesthetizing and Dissection: Rabbits were anesthetized using chloroform and then dissected. Organs of interest (liver, kidney) were immediately fixed in 10% formalin for 24 hours for histological examination. Histological Section Preparation: Tissue sections were prepared following standard histological techniques, including dehydrating, clearing, and embedding in paraffin, before being sectioned and stained for microscopic examination. Statistical Analysis: Data were analyzed using the t-test and ANOVA, with comparisons of mean values made using the Dunnett's multiple comparison test, setting the significance level at p ≤ 0.05 [17, 18].
These modifications aim to clarify the methodology, ensuring reproducibility and understanding of the experimental procedures.
The results presented in Table 1 indicate a significant elevation (P<0.05) in the serum Glucose concentration in both the C. krusei group and C. krusei + Nystatin cluster related to the mechanism group. Conversely, there was a noteworthy reduction in the Chitosan cluster related to the mechanism group, and no significant decrease was observed in the Chitosan cluster related to the controller group. Notably, important differences were observed in the C. krusei + Chitosan collection associated to the controller group.
Table 1. Chitosan treatment's effects on a few rabbits’ biochemical tests
|
Parameters Groups |
Glucose (mg/dl) |
Uric Acid (mg/dl) |
Urea (mg/dl) |
Creatinine (mg/dl) |
|
Control |
107.7±1.80 bc |
3.80± 0.10 a |
37.00±2.12 Ab |
0.400±0.08 b |
|
Chitosan |
88.00± 1.26 d |
3.30± 0.11 b |
30.20± 2.79 C |
0.340±0.09 b |
|
C. krusei |
119.0± 1.00 a |
3.89± 0.21 a |
40.04± 2.11 A |
0.800±0.100 a |
|
C. krusei+Chitosan |
111.7± 1.08 b |
3.71± 0.13 a |
36.10 ± 2.01 B |
0.710±0.103 a |
|
C. krusei+Nystatin |
117.2± 2.16 a |
3.90± 0.15 a |
39.00± 2.22 A |
0.597±0.181 a |
-The numbers stand for mean ±S.E.
-Vertically nasty significant variance with a variety of letters at the significance level (P<0.05).
Figure 1. Chitosan treatment's effects on a few rabbits’ biochemical tests
The findings in Table 1 indicate an important decrease (P<0.050) in the serum concentrations of Uric acid and Urea in the Chitosan cluster associated to the controller group. However, there were no important alterations in the C. krusei group, C. krusei + Chitosan cluster, and C. krusei + Nystatin assembly related to the controller group. Also the Figure 1 shows the decrease in the serum concentration as a chart to explication the effect of chitosan treatment between all groups.
The outcomes in Table 1 revealed a significant rise (P<0.050) in the concentration of Creatinine in the plasma in the C. krusei cluster, C. krusei + Chitosan cluster, and C. krusei + Nystatin cluster associated to the controller group. Meanwhile, there were no significant alterations in the Chitosan cluster associated to the mechanism group.
3.1 Effects on glucose metabolism
The observed increase in serum glucose levels in the C. krusei and C. krusei + Nystatin groups contrasts with the noteworthy reduction in the Chitosan group [19]. This finding underscore chitosan's potential role in modulating glucose metabolism, possibly through inhibition of lipid oxidation, RNA breakdown, and proinsulin inhibition mechanisms that are commonly activated by active oxygen species [20, 21]. Such mechanisms contribute to the stabilization of serum glucose by affecting glycogenolysis in the liver and muscles. Our observations align with existing literature that documents the glucose-lowering effects of chitosan, highlighting its potential as a therapeutic agent in conditions characterized by impaired glucose metabolism [22].
3.2 Renal function markers
The elevated levels of creatinine, uric acid, and urea in the serum upon treatment with C. krusei suggest a compromised renal filtration rate [23], indicating the nephrotoxic potential of C. krusei infection. This decrease in renal function contrasts with the protective effects observed in the Chitosan group, where these markers were significantly reduced, indicating an improvement in the kidneys' ability to excrete wastes [24]. The protective role of chitosan against renal damage supports its potential utility in mitigating infection-induced nephrotoxicity, a significant finding given the scarcity of literature on chitosan's renal protective effects [25].
3.3 Serum protein profiles
As shown in Table 2, a significant increase in the total protein absorption in the serum was noted in the Chitosan group compared to the mechanism cluster, while a decrease was observed in the C. krusei group. Similarly, albumin levels were notably reduced in the C. krusei group but remained stable or improved in groups treated with chitosan [26, 27]. The increase in globulin levels in the Chitosan-treated groups suggests an enhanced immune response or an improvement in overall protein synthesis [22, 23]. These changes in protein profiles not only reflect chitosan's potential impact on protein metabolism but also suggest its role in supporting immune function during infections.
3.4 Clinical relevance and safety implications
The clinical relevance of our findings lies in chitosan's potential as a novel therapeutic agent for managing fungal infections, particularly those caused by C. krusei, a pathogen known for its resistance to many antifungal drugs. The implications for patient safety are significant, considering the observed benefits of chitosan in modulating glucose metabolism, improving renal function, and potentially enhancing immune response without adverse effects on liver and kidney histology. These benefits suggest that chitosan could offer a safe, effective alternative to traditional antifungal treatments, emphasizing the need for further clinical studies to fully understand its therapeutic potential and safety profile [28, 29].
Table 2. Influence of chitosan treatment on rabbit serum total protein
|
Parameters Groups |
Total Protein (g/dl) |
Albumine (g/dl) |
Globulin (g/dl) |
|
Control |
6.00±0.13 B |
3.50±0.10 a |
2.50±0.07 B |
|
Chitosan |
6.61±0.61 A |
3.80±0.13 a |
2.81±0.08 A |
|
C. krusei |
5.00±0.50 C |
3.00±0.11 b |
2.00±0.10 C |
|
C. krusei+Chitosan |
5.90±0.70 B |
3.30±0.14 b |
2.60±0.11 A |
|
C. krusei+Nystatin |
5.59±0.92 B |
2.90±0.09 c |
2.69±0.06 A |
-The numbers stand for mean ±S.E.
-Vertically nasty significant variation in letter dissimilarity at the significance level (P<0.050).
Figure 2. Influence of chitosan behavior on rabbit serum total protein
Figure 3. The lipid profiles of rabbits treated with chitosan
The reduction in total protein as shown in Figure 2, albumin, and globulin levels observed in C. krusei infections represents a critical disruption in protein synthesis, potentially leading to significant damage across various internal organs, most notably within liver cells [22]. This observation is particularly concerning given the liver's crucial role in detoxification and protein synthesis. Fungal infections, especially those caused by Candida species, are a recognized complication in patients undergoing chemotherapy for malignant blood tumors, where the gastrointestinal tract often serves as a reservoir for Candida proliferation [23].
Our results, as presented in Table 3 and Figure 3, showed a significant improvement in lipid profile markers in the Chitosan-treated groups compared to the control group. This includes a notable reduction in cholesterol and triglyceride levels [30], and an increase in high-density lipoprotein (HDL) levels within the serum of the Chitosan, C. krusei + Chitosan, and C. krusei + Nystatin groups. Such lipid-modulating effects of chitosan were not observed in the C. krusei alone and C. krusei + Nystatin groups, highlighting chitosan's potential role in managing dyslipidemia associated with fungal infections [24].
Contrastingly, low-density lipoprotein (LDL) and very low-density lipoprotein (VLDL) levels saw a significant decrease in the serum of the Chitosan group compared to the control [31], suggesting that chitosan might also contribute to the reduction of cardiovascular risk factors. No significant differences in these lipid markers were noted in the C. krusei, C. krusei + Chitosan, and C. krusei + Nystatin groups when compared to the control group, further underscoring the specific lipid-modulating effect of chitosan in this experimental setup [25].
These findings imply that beyond its potential antifungal properties, chitosan could offer therapeutic benefits in correcting lipid imbalances induced by C. krusei infections or possibly by other fungal pathogens [32]. The clinical relevance of these observations lies in chitosan's potential as a complementary treatment option, not only in combating fungal infections but also in managing associated metabolic disruptions, thus enhancing patient care in individuals at risk of or suffering from fungal infections [33].
Table 3. Chitosan treatment's effects on rabbits' lipid profiles
|
Parameters Groups |
Cholestrol mg/dl |
Triglyceride mg/dl |
HDL mg/dl |
LDL mg/dl |
VLDL mg/dl |
|
Control |
157.0±2.50 a |
120.0±2.11 a |
31.40±1.23 b |
101.6±3.23 A |
24.00±1.00 a |
|
Chitosan |
143.0±2.14 c |
110.0±1.20 b |
35.00±0.54 a |
86.00±3.27 B |
22.00±0.82 b |
|
C. krusei |
156.5±2.02 a |
117.0±3.26 a |
29.2±0.90 b |
103.90±5.30 A |
23.40±1.11 a |
|
C. krusei+Chitosan |
151.0±2.10 b |
113.0±2.28 b |
30.30±1.23 b |
98.10±3.90 A |
22.50±0.76 ab |
|
C. krusei+Nystatin |
153.0±2.21 b |
119.0±3.21 a |
30.60±1.00 b |
98.70±4.21 A |
23.80±0.59 a |
-The numbers stand for median ± S.E.
-Variance of distinct characters in a vertical nasty important at the significance level (P<0.050).
3.5 Chitosan's role in modulating fat metabolism
Stress and the occurrence of fat oxidation alongside unsaturated fatty acids may lead to disruptions in fat metabolism. This includes the inhibition of the secretion and excretion of steroids and bile salts, as well as disturbances in the intestinal absorption processes [24]. Additionally, oxidative stress can trigger an increase in the intestinal absorption of cholesterol, primarily through the upregulated activity of the enzyme cholesterol acyl transferase [25], a process potentially exacerbated by insulin deficiency affecting pancreatic beta cells [26].
Chitosan has been shown to mitigate these metabolic disruptions, notably lowering cholesterol, triglycerides, low-density lipoproteins (LDL), and very low-density lipoproteins (VLDL), while simultaneously increasing high-density lipoprotein (HDL) levels [27]. It is postulated that chitosan degrades into polysaccharides within the intestine, leading to an increase in viscosity and a consequent reduction in fat and cholesterol absorption [28]. Furthermore, the transformation of chitosan into glucosamine may reduce triglyceride synthesis in the liver [29].
3.6 Microscopic analysis of liver tissue
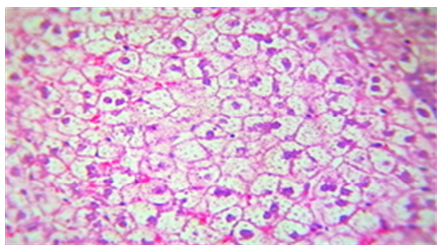
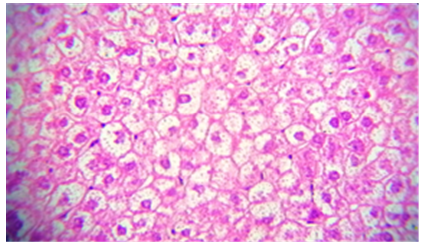
The liver tissue's microscopic examination exposed evidence of hemorrhage and hemolysis, alongside significant inflammatory cell infiltration around the portal vein. A notable presence of macrophages (Kupffer cells) within the sinusoids was also observed. Moreover [33], fatty necrosis was prevalent in a majority of hepatocytes, highlighting the protective and reparative roles of chitosan against liver damage induced by stress factors and possibly by the metabolic disturbances associated with fungal infections, as shown in Figures 4-9, which starkly contrast the findings in the control group.
Figure 4. The histological slice of the chitosan-treated liver revealed: A) hemorrhage within parenchyma as well as B) hemolysis
Figure 5. The histological slice of the chitosan-treated liver revealed: A) Inflammatory lymphatic infiltrate within portal area, B) Sinusoid were filled with kuppfer cells
Figure 6. The histological slice of the liver treated with Nystatin and C. Krusei revealed: steatosis is present in wide areas of liver
Figure 7. Histological slice of the C. krusei-treated liver revealed: steatosis is present in wide areas of liver
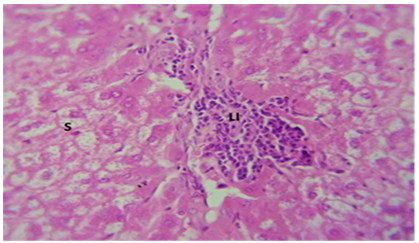
Figure 8. Histological slice of the C. krusei-treated liver revealed: S) steatosis are present with patch of hepatocytes. LI) Lymphocytes infiltration
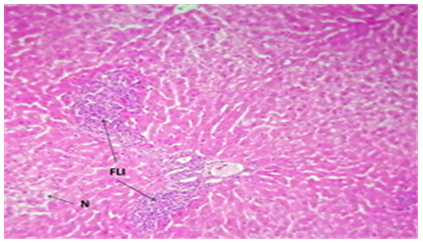
Figure 9. The histological slice of the liver treated with Nystatin and C. Krusei revealed: FLI) focal lymphocytic infiltration in the portal area. N) Some necrotic cells are observed
The liver, with its complex network of enzymatic pathways, plays a crucial role in the body's physiological functions, making it particularly susceptible to alterations by numerous drugs and toxic substances [30]. Our study has revealed extensive hypothermic injuries across all experimental groups, characterized notably by the swelling and degeneration of hepatocytes [34].
The observed swelling and deterioration of hepatocytes are likely outcomes of disruptions in metabolic processes within these cells, influenced by the concentration of various substances in the blood [35]. The disintegration of cell nuclei suggests heightened vulnerability to damage, particularly because hepatocytes are primary agents in disseminating substances through the portal circulation from the gastrointestinal tract [36]. This vulnerability is exacerbated when the cells' nutritional status, exposure to toxins, and the duration of such exposures are compromised, or when drug interactions affect cellular membranes and ionic balances [37].
The widespread damage to hepatocytes, including cytoplasmic degradation and nuclear disintegration, and the induction of apoptosis across most cells, highlights the severe impact of the studied conditions on liver health [38]. The process of programmed cell death (apoptosis) is a natural mechanism for removing excess or damaged cells in tissues, including during certain diseases. It involves a series of steps such as chromatin condensation, DNA fragmentation, and the formation of apoptotic bodies, which are then eliminated by phagocytosis without eliciting an inflammatory response [39, 40].
Furthermore, the microscopic examination of kidney sections revealed significant atrophy of renal glomeruli and necrosis of the cells lining the urinary glomeruli. Notably, there was an expansion of urinary tubules and the presence of necrotic cell debris within the epithelial cells lining the urine tubules and their lumens, as depicted in Figures 10-15, starkly contrasting with observations from the control group shown in Figures 10-17 [41].
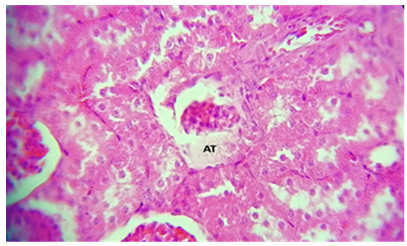
Figure 10. A histological slice of the kidney after chitosan treatment revealed: AT) Atrophy of the glomerulus
Figure 11. A histological slice of the kidney after chitosan treatment revealed: N) necrosis of the glomerulus
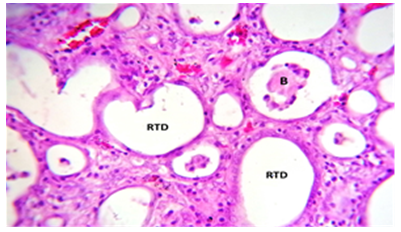
Figure 12. The kidney treated with C. krusei and nystatin had a histological section that revealed: RTD) renal tubules dilation with B) debris of endothelium within its lumen
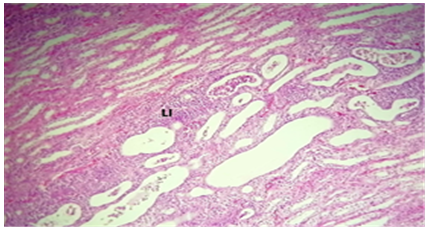
Figure 13. The kidney treated with C. krusei and nystatin had a histological section that revealed: LI) infiltration of lymphocytes intra renal tubules as well as RTD) renal tubules dilation
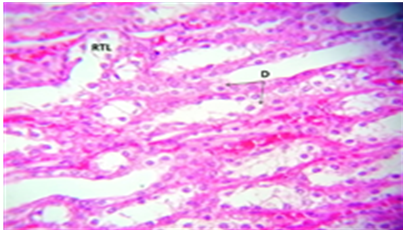
Figure 14. A portion of the kidney treated with C. krusei's histology revealed: RTL) renal tubules Laceration. D) Degeneration of renal tubules endothelial cells
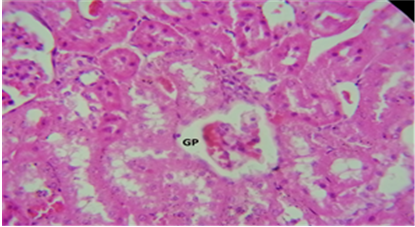
Figure 15. The kidney treated with C. krusei + Chitosan displayed a histological section that revealed: GP) glomerulus prolapse and begin with atrophy
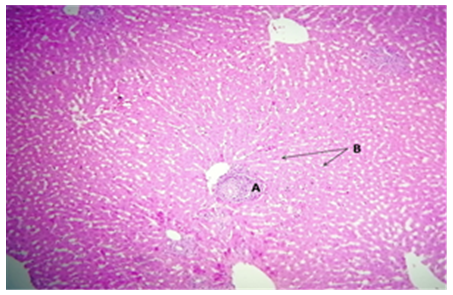
Figure 16. The control group's rabbit liver had a standard histological construction for the surrounding hepatocytes (HC) and central vein (CV)
Figure 17. Rabbit kidney in the mechanism cluster: glomeruli (G) and surrounding tubules (T) in the cortical section exhibit normal histological structure
This study highlighted that the kidney is notably susceptible to the effects of Candida yeast and chitosan [42], manifesting a range of damage from necrosis in urinary tubules and glomeruli to varying degrees, including complete damage, alongside vascular injury and significant tissue congestion. These observations suggest that such impacts may arise from alterations in mineral homeostasis and potential adverse effects of therapeutic agents [43]. The occurrence of urinary tubular necrosis, often attributed to toxicity, can stem from the deprivation of oxygen to epithelial cells lining the urinary tubules, crucially dependent on adequate blood supply for their metabolic functions. Damage to blood vessels, whether through necrosis or constriction, can significantly impair blood flow, leading to oxygen shortages and subsequent cellular distress [44, 45].
The histological examination revealed enlargement and swelling of cells within the renal tubules, leading to tubular obstruction [46]. This aligns with findings that toxic substances can induce tubular cell swelling, thereby causing blockages within the tubular lumen and cell necrosis. Such effects result from disrupted sodium balance [47], leading to fluid retention and swelling, and oxygen deficiency, hindering mitochondrial energy production [48]. Furthermore, the presence of a homogeneous pink amyloid fluid in tissue samples points to systemic amyloidosis, where acidophilic extracellular protein deposits can inflict damage on surrounding tissues [49]. This condition underscores the systemic impact of renal tubular and vascular damage induced by toxic exposure and metabolic imbalances [50, 51].
These findings underscore the critical need for cautious management of treatments involving Candida infections and highlight the importance of safeguarding renal health against the multifaceted damage mechanisms induced by infections and therapeutic interventions.
Our findings suggest that chitosan contributes to improvements in kidney function, as evidenced by reductions in the levels of urea, uric acid, and glucose in the blood. This clarification aims to precisely specify the kidney function parameters that were observed to improve, thereby addressing the ambiguous term previously referred to as "ureic urea."
We observed an association between chitosan treatment and an increase in serum total protein, including albumin and globulin levels. In contrast, the presence of C. krusei was associated with an alteration in these serum proteins. It is to be emphasized that these findings imply relations but not causal effects. In relation to lipid metabolism, our study identified associations with chitosan treatment and reduced cholesterol, triglycerides, LDL, and VLDL levels in the circulation. This phrasing has been purposefully selected to mirror the seen connections but avoiding direct inferences.
C. krusei was harmful to liver and kidney tissues. The conclusion is derived straight from the data provided and emphasizes the necessity of further study on the pathogenic impact of C. krusei on these organs. While our study points to the potential benefits of chitosan in renal function enhancement and lipid profile modulation, we approach the clinical implications of these findings with caution. The therapeutic potential of chitosan, particularly in the context of renal function and lipid metabolism, warrants further research. Comprehensive clinical trials are necessary to fully elucidate the efficacy, safety, and mechanisms by which chitosan exerts its effects, prior to making any clinical recommendations.
[1] Adeva-Andany, M.M., González-Lucán, M., Donapetry-García, C., Fernández-Fernández, C., Ameneiros-Rodríguez, E. (2016). Glycogen metabolism in humans. BBA Clinical, 5: 85-100. https://doi.org/10.1016/j.bbacli.2016.02.001
[2] Abdelal, W. (2012). From the Mosque to Satellite Broadcasting: A Historical Perspective of Hamas Media Strategy. University of Exeter, United Kingdom.
[3] Allen, T., Allen, T.D., Cowling, G. (2011). The Cell: A Very Short Introduction. Oxford University Press, USA.
[4] Bakshi, P.S., Selvakumar, D., Kadirvelu, K., Kumar, N.S. (2020). Chitosan as an environment friendly biomaterial–A review on recent modifications and applications. International Journal of Biological Macromolecules, 150: 1072-1083. https://doi.org/10.1016/j.ijbiomac.2019.10.113
[5] Behmagham, F., Mustafa, M.A., Saraswat, S.K., Khalaf, K.A., Kaur, M., Ghildiyal, P., Vessally, E. (2024). Recent investigations into deborylative (thio-/seleno-) cyanation of aryl boronic acids. RSC Advances, 14(13): 9184-9199. https://doi.org/10.1039/D4RA00487F
[6] Bellavia, M. (2018). Evaluation of targeting schemes for complement-modulating, strain-selective antimicrobial microparticles. Doctoral Dissertation. Georgia Institute of Technology.
[7] Beth, D., Robert, G.T. (2004). Basic & Clinical Biostatistics 4/E (EBOOK). McGraw-Hill Medical Publishing Division, New York.
[8] Brack, W., Escher, B.I., Müller, E., Schmitt-Jansen, M., Schulze, T., Slobodnik, J., Hollert, H. (2018). Towards a holistic and solution-oriented monitoring of chemical status of European water bodies: How to support the EU strategy for a non-toxic environment? Environmental Sciences Europe, 30: 1-11. https://doi.org/10.1186/s12302-018-0161-1
[9] Brunzel, N. A. (2021). Fundamentals of Urine and Body Fluid Analysis-E-Book. Elsevier Health Sciences.
[10] Buzea, C., Pacheco, I.I., Robbie, K. (2007). Nanomaterials and nanoparticles: Sources and toxicity. Biointerphases, 2(4): MR17-MR71. https://doi.org/10.1116/1.2815690
[11] Campos, V.C., Tappy, L. (2016). Physiological handling of dietary fructose-containing sugars: Implications for health. International Journal of Obesity, 40(1): S6-S11. https://doi.org/10.1038/ijo.2016.8
[12] Chavez-Tapia, N.C., Lizardi-Cervera, J., Perez-Bautista, O., Ramos-Ostos, M.H., Uribe, M. (2006). Smoking is not associated with nonalcoholic fatty liver disease. World Journal of Gastroenterology: WJG, 12(32): 5196-5200. https://doi.org/10.3748%2Fwjg.v12.i32.5196
[13] Chowdhary, H., Chaudhary, D.N.K., Harahsheh, F.A.H., Mustafa, M.A., Rajak, D.M., Tomar, R.K. (2024). Technical analysis of internet shutdowns: Economic and cybersecurity dimensions in India and international context. Journal of Theoretical and Applied Information Technology, 102(4): 1588-1603.
[14] Dabla, P.K. (2010). Renal function in diabetic nephropathy. World Journal of Diabetes, 1(2): 48-56. https://doi.org/10.4239%2Fwjd.v1.i2.48
[15] Deng, X., Ye, Z., Cao, H., Bai, Y., Che, Q., Guo, J., Su, Z. (2020). Chitosan oligosaccharide ameliorated obesity by reducing endoplasmic reticulum stress in diet-induced obese rats. Food & Function, 11(7): 6285-6296. https://doi.org/10.1039/D0FO01107J
[16] Fiore, M., Cascella, M., Bimonte, S., Maraolo, A.E., Gentile, I., Schiavone, V., Pace, M.C. (2018). Liver fungal infections: An overview of the etiology and epidemiology in patients affected or not affected by oncohematologic malignancies. Infection and Drug Resistance, 11: 177-186. https://doi.org/10.2147/IDR.S152473
[17] Gaim, K., Gebru, G., Abba, S. (2015). The effect of arsenic on liver tissue of experimental animals (fishes and mice)—A review article. International Journal of Scientific and Research Publications, 5(5): 1-9. https://doi.org/10.1007/978-3-319-24584-3_55
[18] Gerber, P.A., Rutter, G.A. (2017). The role of oxidative stress and hypoxia in pancreatic beta-cell dysfunction in diabetes mellitus. Antioxidants & Redox Signaling, 26(10): 501-518. https://doi.org/10.1089/ars.2016.6755
[19] Giglio, R.V., Carruba, G., Cicero, A.F., et al. (2020). Pasta supplemented with Opuntia ficus-indica extract improves metabolic parameters and reduces atherogenic small dense low-density lipoproteins in patients with risk factors for the metabolic syndrome: A four-week intervention study. Metabolites, 10(11): 428. https://doi.org/10.3390/metabo10110428
[20] Gomaraschi, M., Bonacina, F., Norata, G.D. (2019). Lysosomal acid lipase: From cellular lipid handler to immunometabolic target. Trends in Pharmacological Sciences, 40(2): 104-115. https://doi.org/10.1016/j.tips.2018.12.006
[21] Hsu, C.Y., Mustafa, M.A., Yadav, A., et al. (2024). N2 reduction to NH3 on surfaces of Co-Al18P18, Ni-Al21N21, Fe-B24N24, Mn-B27P27, Ti-C60 and Cu-Si72 catalysts. Journal of Molecular Modeling, 30(3): 1-11. https://doi.org/10.1007/s00894-024-05862-y
[22] Hussain, T., Tan, B., Murtaza, G., et al. (2020). Flavonoids and type 2 diabetes: Evidence of efficacy in clinical and animal studies and delivery strategies to enhance their therapeutic efficacy. Pharmacological Research, 152: 104629. https://doi.org/10.1016/j.phrs.2020.104629
[23] Ibrahim, S.R., Mohamed, G.A., Banjar, Z.M., Kamal, H.K. (2013). Natural antihyperlipidemic agents: Current status and future perspectives. Phytopharmacology, 4(3): 492-531.
[24] Jamiu, A.T., Albertyn, J., Sebolai, O.M., Pohl, C.H. (2021). Update on Candida krusei, a potential multidrug-resistant pathogen. Medical Mycology, 59(1): 14-30. https://doi.org/10.1093/mmy/myaa031
[25] Khan, A.A., Alanazi, A.M., Alsaif, N., Algrain, N., Wani, T.A., Bhat, M.A. (2021). Enhanced efficacy of thiosemicarbazone derivative-encapsulated fibrin liposomes against candidiasis in murine model. Pharmaceutics, 13(3): 333. https://doi.org/10.3390/pharmaceutics13030333
[26] Kumar, C.R. (2003). Basic Pathology. 7th ed. SAUNDERS, Philadelaphia.
[27] Lo, W.H., Deng, F.S., Chang, C.J., Lin, C.H. (2020). Synergistic antifungal activity of chitosan with fluconazole against Candida albicans, Candida tropicalis, and fluconazole-resistant strains. Molecules, 25(21): 5114. https://doi.org/10.3390/molecules25215114
[28] Mejía, N., Mustafa, M.A., Kumar, A., et al. (2024). Potential of nanocages as effective catalysts for oxygen reduction reaction. Silicon, 1-8. https://doi.org/10.1007/s12633-024-02944-7
[29] Morrissey, S.E., Newth, T., Rees, R., Barr, A., Shora, F., Laycock, J.F. (2001). Renal effects of recombinant prolactin in anaesthetized rats. European Journal of Endocrinology, 145(1): 65-71. https://doi.org/10.1530/eje.0.1450065
[30] Mudgil, D. (2017). The interaction between insoluble and soluble fiber. In: Dietary Fiber for the Prevention of Cardiovascular Disease, pp. 35-59.
[31] Ogbe, R.J., Ochalefu, D.O., Mafulul, S.G., Olaniru, O.B. (2015). A review on dietary phytosterols: Their occurrence, metabolism and health benefits. Asian Journal of Plant Science and Research, 5(4): 10-21.
[32] Ortiz, D.T.C., Ghadir, G.K., Mustafa, M.A., Chandra, S., Kaur, I., Saadh, M.J., Alawaideh, Y.M., Elawady, A., Elmasry, Y. (2024). Exploring the photovoltaic performance of boron carbide quantum dots doped with heteroatoms: A DFT analysis. Diamond and Related Materials, 143: 110933. https://doi.org/10.1016/j.diamond.2024.110933
[33] Pais, G.M., Liu, J., Zepcan, S., Avedissian, S.N., Rhodes, N.J., Downes, K.J., Moorthy, G.S., Scheetz, M.H. (2020). Vancomycin‐induced kidney injury: Animal models of toxicodynamics, mechanisms of injury, human translation, and potential strategies for prevention. Pharmacotherapy: The Journal of Human Pharmacology and Drug Therapy, 40(5): 438-454. https://doi.org/10.1002/phar.2388
[34] Ridley, J.W. (2018). Metabolic origins of urine and other body fluids. In: Fundamentals of the Study of Urine and Body Fluids, pp. 45-60. https://doi.org/10.1007/978-3-319-78417-5_4
[35] Saadh, M.J., Avecilla, F.R.B., Mustafa, M.A., Kumar, A., Kaur, I., Alawayde, Y.M., Mahmud, S.F., Sattar, R., Elmasry, Y. (2024). The promising role of doped h-BANDs for solar cells application: A DFT study. Journal of Photochemistry and Photobiology A: Chemistry, 451: 115499. https://doi.org/10.1016/j.jphotochem.2024.115499
[36] Saadh, M.J., Lagum, A.A., Ajaj, Y., Saraswat, S.K., Dawood, A.A.A.S., Mustafa, M.A., Alawadi, A., Omran, A.A., Elmasry, Y. (2024). Adsorption behavior of Rh-doped graphdiyne monolayer towards various gases: A quantum mechanical analysis. Inorganic Chemistry Communications, 160: 111928. https://doi.org/10.1016/j.inoche.2023.111928
[37] Saadh, M.J., Mustafa, M.A., Batoo, K.M., et al. (2024). Performances of nanotubes and nanocages as anodes in Na-ion battery, K-ion battery, and Mg-ion battery. Ionics, 1-8. https://doi.org/10.1007/s11581-024-05440-5
[38] Santos, D.K.C., Mustafa, M.A., Bansal, P., et al. (2024). Investigation of ORR and OER mechanisms by co-and fe-doped silicon nanocages (Si48 and Si60) and Co-and Fe-doped silicon nanotubes (SiNT (5, 0) and SiNT (6, 0)) as Acceptable Catalysts. Silicon, 1-13. https://doi.org/10.1007/s12633-024-02915-y
[39] Skulachev, V.P. (2002). Programmed death phenomena: From organelle to organism. Annals of the New York Academy of Sciences, 959(1): 214-237. https://doi.org/10.1111/j.1749-6632.2002.tb02095.x
[40] Stricker, S.A. (2022). Human Microanatomy: Cell Tissue and Organ Histology with Celebrity Medical Histories. CRC Press.
[41] Sweeney, M.F., Hasan, N., Soto, A.M., Sonnenschein, C. (2015). Environmental endocrine disruptors: Effects on the human male reproductive system. Reviews in Endocrine and Metabolic Disorders, 16(4): 341-357. s://doi.org/10.1007/s11154-016-9337-4
[42] Valverde, V., Ortiz, D.T.C., Mustafa, M.A., Kumar, A., Kaur, I., Karim, M.M., Al-Khamiseh, B.M., Elawady, A., Lasisi, A. (2024). Design gas sensor based on transition metal doped graphene like nanosheets: A quantum chemical study. Diamond and Related Materials, 143: 110895. https://doi.org/10.1016/j.diamond.2024.110895
[43] Weska, R.F., Moura, J.M.D., Batista, L.D.M., Rizzi, J., Pinto, L.D.A. (2007). Optimization of deacetylation in the production of chitosan from shrimp wastes: Use of response surface methodology. Journal of Food Engineering, 80(3): 749-753. https://doi.org/10.1016/j.jfoodeng.2006.02.006
[44] Willner, E.L., Tow, B., Buhman, K.K., Wilson, M., Sanan, D.A., Rudel, L.L., Farese Jr, R.V. (2003). Deficiency of acyl CoA: Cholesterol acyltransferase 2 prevents atherosclerosis in apolipoprotein E-deficient mice. In Proceedings of the National Academy of Sciences, 100(3): 1262-1267. https://doi.org/10.1073/pnas.0336398100
[45] Alabbasy, R.H., Azeez, A.K., Meri, M.A., Mustafa, M.A. (2023). Histological study of the effect of some oncology drugs on heart muscle. AIP Conference Proceedings, 2977(1).
[46] Wong Rebecca, S.Y. (2011). Apoptosis in cancer: From pathogenesis to treatment. Journal of Experimental and Clinical Cancer Research, 30: 87. https://doi.org/10.1186/1756-9966-30-87
[47] Xu, W., Mohan, A., Pitts, N.L., Udenigwe, C., Mason, B. (2020). Bile acid-binding capacity of lobster shell-derived chitin, chitosan and chitooligosaccharides. Food Bioscience, 33: 100476. https://doi.org/10.1016/j.fbio.2019.100476
[48] Abdulqader, A.T., Al-Sammarie, A.M.Y., Mustafa, M.A. (2022). A comparative environmental study of aqueous extracts of ginger and grapes to protect hepatocytes in Albino rabbits and a comparison of extracts in preserving Awassi lamb meat from oxidation. IOP Conference Series: Earth and Environmental Science, 1029(1): 012001.
[49] Khaleel, Z.I., Saab, N.G., Meri, M.A., Mustafa, M.A. (2023). The role of microbial pathogens in infection of lung organs and spleen of laboratory albino rats. AIP Conference Proceedings, 2977(1).
[50] Zhang, Y., Chen, X., Gueydan, C., Han, J. (2018). Plasma membrane changes during programmed cell deaths. Cell Research, 28(1): 9-21. https://doi.org/10.1038/cr.2017.133
[51] Zheng, H., Jiang, J., Xu, S., Liu, W., Xie, Q., Cai, X., Zhang, J., Liu, S., Li, R. (2021). Nanoparticle-induced ferroptosis: Detection methods, mechanisms and applications. Nanoscale, 13(4): 2266-2285. https://doi.org/10.1039/D0NR08478F