Antibacterial and Anticancer Activities of Bioactive Compounds Produced by Bacillus thermoamylovorans

Manal K. Mohammad*![]() | Suzan S. Hussain
| Suzan S. Hussain![]() | Amer S. Mahdi
| Amer S. Mahdi![]()
© 2025 The authors. This article is published by IIETA and is licensed under the CC BY 4.0 license (http://creativecommons.org/licenses/by/4.0/).
OPEN ACCESS
The antibacterial and anticancer potential of Bacillus thermoamylovorans isolates was evaluated, and demonstrated activity against the tested uropathogenic strains such as Escherichia coli, Staphylococcus aureus, Pseudomonas aeruginosa, and Klebsiella pneumoniae. The maximum production of bioactive compounds was achieved when B. thermoamylovorans was cultured in tryptic soy broth supplemented with 40% glucose, adjusted to pH 7, and incubated at 50℃ for 96 hours. Further evaluation of minimum inhibitory concentration (MIC) and minimum bactericidal concentration (MBC) for selected bioactive compounds (cis-vaccenic acid, linoleic acid, oleic acid, dibutyl phthalate, diethyl phthalate, and ascorbic acid) demonstrated that cis-vaccenic acid exhibited the strongest activity, with values of MIC from 32 to128 µg/mL, and values of MBC from 64-256 µg/mL, particularly effective against S. aureus and K. pneumoniae. Diethyl phthalate showed moderate bactericidal potential (MBC 256-1024 µg/mL), while other compounds required higher concentrations (512-2048 µg/mL). P. aeruginosa showed strong resistance to most compounds, except for cis-vaccenic acid (MIC 64 µg/mL, MBC 128 µg/mL) and oleic acid (MIC 256 µg/mL). Cytotoxicity assays (MTT) against MCF-7 and HeLa cancer cell lines revealed potent effects of purified cis-vaccenic acid (IC₅₀ = 20 µg/mL for MCF-7, 12.9 µg/mL for HeLa), compared to diethyl phthalate (IC₅₀ = 445 µg/mL for MCF-7, 369 µg/mL for HeLa). These findings highlight the promising antibacterial and cytotoxicity potential of bioactive metabolites derived from B. thermoamylovorans.
antimicrobial, anticancer, Bacillus thermoamylovorans, cis-vaccenic acid, diethyl phthalate
The genus Bacillus, belonging to the phylum Bacillota, consists of rod-shaped, Gram-positive bacteria capable of forming endospores that confer resistance to harsh environmental conditions such as desiccation, heat, and chemical stress. These bacteria can grow under aerobic or facultatively anaerobic conditions and are widely distributed in soil, water, and as part of the human and animal microbiota. Owing to their ability to produce antibiotics, enzymes, and other bioactive metabolites, Bacillus species play essential roles in medicine, agriculture, and industry. Molecular studies, particularly 16S rRNA sequencing, have revealed significant taxonomic revisions within this genus, resulting in the reclassification of several species into related genera such as Paenibacillus, Aneurinibacillus, and Brevibacillus [1, 2]. Among the different members of the Bacillus genus, thermophilic species have gained increasing attention due to their ability to produce bioactive metabolites and thermostable enzymes with medical and industrial applications. Within this group, particularly Bacillus thermoamylovorans is noteworthy because of its high amylolytic capacity, thermophilic nature, and potential to synthesize heat-stable metabolites. These characteristics make it a promising candidate for biotechnological exploration, compared with other Bacillus species.
Bacillus thermoamylovorans, first described in 1995, is a thermophilic facultative anaerobe capable of producing heat-resistant spores and growing optimally at 50℃. Its name reflects the capacity of utilizing starch at elevated temperatures. Colonies are typically circular, white, and 2–3 mm in diameter, with motility conferred by flagella. The organism metabolizes carbohydrates such as lactose and glucose, making it a potential candidate for biofuel production through the degradation of organic materials from sources such as sewage sludge [3, 4].
During the stationary growth phase, Bacillus species produce secondary metabolites—organic compounds not directly involved in growth or reproduction but vital for ecological functions such as defense, competition, and signaling. These metabolites include polyketides, lipopeptides (e.g., surfactin), and antimicrobial peptides. Many of them display potent antibacterial, antioxidant, and antiviral activities, disrupting bacterial membranes, mitigating oxidative stress-related diseases, and interfering with viral replication [5, 6]. Owing to this unique chemical diversity, Bacillus metabolites represent a promising source of new pharmaceutical agents, particularly against antibiotic-resistant pathogens. Specifically, B. thermoamylovorans offers potential for medical and biotechnological applications, and optimizing its biosynthetic pathways could unlock novel therapeutic opportunities.
Despite the growing interest in Bacillus species as producers of bioactive metabolites, limited studies have focused on B. thermoamylovorans. Most available research has highlighted its thermophilic nature and industrial potential, whereas its capacity to synthesize antimicrobial and anticancer compounds remains poorly investigated [7]. Although many species of thermophilic Bacillus have been reported to produce antibacterial peptides, thermostable enzymes, and different bioactive compounds, specifically the literature describing the secondary metabolites of Bacillus thermoamylovorans is still scarce. On this species, previous studies have focused on its heat-resistant spores primarily, industrial applications in starch and biofuel production, and amylolytic activity degradation. Have identified potential bioactive molecules in only a few reports, like thermostable amylases and antibacterial compounds, but these findings lack detailed biological evaluation and chemical characterization. Therefore, B. thermoamylovorans may produce metabolites with antibacterial or therapeutic potential despite preliminary evidence suggesting that comprehensive studies on their biological activity and composition remain limited—highlighting a clear research gap that this study aims to address [8, 9].
Given the alarming rise of antibiotic resistance among clinical pathogens and the global need for safer alternatives in cancer therapy, exploring the secondary metabolites of B. thermoamylovorans may uncover novel compounds with therapeutic relevance. This study, therefore, addresses an important gap by combining microbial isolation, optimized cultivation, chemical characterization, and biological evaluation of the metabolites produced by this species. Cultivation of B. thermoamylovorans in tryptic soy broth at 50℃ under optimized conditions has been reported as effective for bioactive metabolite production [10].
Although limited evidence exists regarding the anticancer potential of B. thermoamylovorans secondary metabolites, some studies have suggested associations with tumor progression under certain conditions [11, 12]. Further research is required to evaluate whether, under optimized growth or genetic modification, this bacterium can yield metabolites with anticancer activities.
From a clinical perspective, urinary tract infections (UTIs) represent one of the most prevalent infections in primary healthcare, often caused by pathogens such as Klebsiella pneumoniae, Escherichia coli, Staphylococcus aureus, and Enterococcus faecalis [13]. Increasing antimicrobial resistance among these uropathogens, largely due to antibiotic overuse, poses a major therapeutic challenge and economic burden [14]. Likewise, breast and cervical cancers remain among the most common malignancies worldwide, with high incidence and mortality rates, emphasizing the need for novel therapeutic agents [15]. In this context, breast cancer (MCF-7) and cervical cancer (HeLa) cell lines are widely used in biomedical research to study molecular mechanisms of carcinogenesis, drug responses, and resistance patterns, as well as to screen for novel anticancer agents [16]. On our previous discovery based on research [8], Bacillus thermoamylovorans produces many metabolites, involving cis-vacenic acid (CVA) and diethyl phthalate (DEP). To systematically evaluate the biological activities of these compounds, we purchased high-purity standards for standardized antibacterial and anticancer activity assessments.
Therefore, the aim of the study is to isolate and identify bioactive metabolites by Bacillus thermoamylovorans, and to evaluate their antibacterial activity against relevant uropathogens clinically, as well as their cytotoxic effects against two cancer cell lines of humans (MCF-7 and HeLa).
2.1 Bacterial strain
The strain of Bacillus thermoamylovorans used in this study was isolated previously and characterized in an earlier publication [8]. In brief, the strain was obtained from the skin of healthy individuals and identified by 16S rRNA sequencing. High similarity to the B. thermoamylovorans sequence. The strain on nutrient agar slants was maintained at 4℃ and subcultured monthly.
2.2 Bioactive compounds
All tested compounds, including linoleic acid, oleic acid, ascorbic acid, diethyl phthalate, dibutyl phthalate, and cis-vaccenic acid, in pure form (≥ 98% purity), were obtained from reputable commercial suppliers. Based on our previous findings, these compounds were selected [8], which showed that Bacillus thermoamylovorans produces different metabolites of similar structure. Using commercially purchased standards ensured consistency, reproducibility, and accuracy in evaluations activity of antibacterial and anticancer [8].
2.3 Antibacterial activity assays (MIC and MBC)
For selected compounds, the antibacterial potential was evaluated against clinically relevant uropathogenic bacterial isolates using minimum inhibitory concentration (MIC) and minimum bactericidal concentration (MBC) assays [17]. For each compound, stock solutions were prepared in 2% dimethyl sulfoxide (DMSO) at concentrations of 2048 µg/mL. In 96-well microtiter plates, broth microdilution was performed containing Mueller–Hinton broth 150 µL per well. To achieve the desired concentration range, serial two-fold dilutions were prepared.
Each well with 20 µL was inoculated of standardized bacterial suspension (1.5 × 10⁸ CFU/mL, McFarland 0.5). The negative control wells contained only sterile broth without inoculum. Plates at 37℃ for 18–20 h were incubated, followed by the addition of resazurin dye 20 µL and further incubation for 2 h. The change of color from blue to pink indicated growth of bacteria; no color change indicated inhibition. MIC was defined as the minimum concentration at which no visible color change was shown. MBC by subculturing from wells was determined with no visible growth onto fresh agar plates to confirm bactericidal activity.
All MIC/MBC assays in triplicate were performed (three technical replicates per concentration) and repeated three times independently.
2.4 Cytotoxicity assays (MTT)
2.4.1 Cell culture
HeLa cells, originally derived from cervical cancer tissue, are characterized by their ability to proliferate indefinitely, making them a well-established model in cancer biology, virology, and vaccine research. MCF-7 cells, derived from human breast carcinoma, express both estrogen and progesterone receptors, and are widely employed as a primary model for investigating breast cancer biology, treatment resistance mechanisms, and drug efficacy.
Human breast cancer (MCF-7) and cervical cancer (HeLa) cells in complete RPMI medium (containing glucose, amino acids, L-glutamine, vitamins, and sodium bicarbonate) supplemented with 1% penicillin–streptomycin and 10% FBS, were grown until confluent monolayers were obtained [18]. With PBS Cells were washed, using trypsin/versene solution detached, in fresh medium resuspended, and seeded into flasks or multi-well plates. Cell counts using a hemocytometer were determined.
2.4.2 Cell treatment
Compounds: Cis-vaccenic acid (CVA; SLBX7301, USA) at 100 mg/mL and diethyl phthalate at 1.12 g/mL (Shanghai Macklin Biochemical Co., Ltd., China) as stock solutions were prepared. These stock solutions in DMSO were subsequently diluted to obtain the working concentrations (0–1000 µg/mL) used in the cell treatment assays.
Cell culture: MCF-7 and HeLa cells were grown in complete RPMI medium until confluent monolayers were obtained. The protocol included:
1. The growth medium was aspirated, and the cells were gently washed with phosphate-buffered saline (PBS).
2. Cells were detached by adding 2–3 mL of trypsin–versene solution and incubating them at 37℃ for 1–2 minutes.
3. Detached cells were suspended in 15–20 ml of fresh RPMI medium and dispersed by pipetting.
4. Cells at the desired concentration were seeded into flasks or multi-well plates and incubated at 37℃ with 5% CO₂.
5. Concentration of cells was determined using a hemocytometer, applying the formula:
Total cell count (cells/mL) = Cell count× Dilution factor × 104
Cells (5,000–6,000 cells/well) in 96-well plates were seeded and incubated for 24 h at 37℃ with 5% CO₂ to allow attachment. After incubation, the medium was replaced with fresh RPMI medium containing the test compounds at the following concentrations:
Compound Concentration Range (µg/mL):
Cis-vaccenic acid: 0, 1, 5, 10, 25, 50, 100
Diethyl phthalate: 0, 1, 5, 10, 25, 50, 100, 250, 500, 1000
Cells were treated for 48 h at 37℃ with 5% CO₂.
2.4.3 MTT assay
After treatment, MTT solution 20 µL (0.5 mg/mL in PBS) was added to each well and incubated for 3–4 h at 37℃. The medium was removed, and 100 µL DMSO was added to dissolve formazan crystals. Absorbance was measured at 570 nm. Cell viability (%) was calculated as:
Cell viability $(\%)=\frac{A_{\text {treatment }}-A_{\text {blank }}}{A_{\text {control }}-A_{\text {blank }}} \times 100$
where, A = absorbance, Negative control with serum-free medium, cells treated, and absorbance of the blank (culture medium and reagent without cells).
All MTT assays in triplicate were performed and independently repeated three times.
2.4.4 IC₅₀ calculation
IC₅₀ values using nonlinear regression were determined with log(inhibitor) vs. response curve fitting in GraphPad Prism 9.
2.5 Statistical analysis
Mean ± standard error (SE) data are expressed. Statistical significance using one-way analysis of variance (ANOVA) was analyzed. Values of p > 0.05 were considered non-significant, while p ≤ 0.05, < 0.01, and < 0.001 were considered significant, highly significant, and very highly significant, respectively.
3.1 Medical applications of bioactive compounds
3.1.1 Antibacterial activity of selected bioactive compounds
The antibacterial efficacy of purified compounds derived from Bacillus thermoamylovorans, including cis-vaccenic acid, linoleic acid, oleic acid, dibutyl phthalate, diethyl phthalate, and ascorbic acid, was evaluated on the basis of their minimum inhibitory concentration (MIC) and minimum bactericidal concentration (MBC) values. These compounds were considered the most effective antibiotic candidates obtained from this bacterium, as they were produced with higher frequency and concentration.
Table 1. MIC of selected bioactive compounds (µg/mL) against uropathogenic isolates (mean ± SD, n = 3)
|
Bacterial Isolate |
Oleic Acid |
Linoleic Acid |
Ascorbic Acid |
Dibutyl Phthalate |
Diethyl Phthalate |
Cis-Vaccenic Acid |
P-value |
|
Escherichia coli |
512 ± 12.7 |
512 ± 12.7 |
512 ± 12.7 |
512 ± 12.7 |
256 ± 4.5 |
128 ± 2.9 |
0.0001 ** |
|
Klebsiella pneumoniae |
512 ± 12.7 |
256 ± 4.5 |
256 ± 4.5 |
256 ± 4.5 |
256 ± 4.5 |
128 ± 2.9 |
0.0001 ** |
|
Pseudomonas aeruginosa |
256 ± 4.5 |
1024±18.7 |
1024±18.7 |
1024±18.7 |
512 ±12.7 |
64 ± 1.04 |
0.0001 ** |
|
Staphylococcus aureus |
256 ± 4.5 |
256 ± 4.5 |
256 ± 4.5 |
512 ± 12.7 |
128 ± 2.9 |
32 ± 0.78 |
0.0001 ** |
Notes: Data as mean ± SD are expressed from three independent experiments. Statistical analysis using the LSD test was performed. P ≤ 0.01 (**) indicates a significant difference.
Figure 1. MIC values of bioactive compounds against uropathogenic isolates
Lower MIC values indicate higher antibacterial potency.
A. Minimum inhibitory concentration (MIC)
MIC assays were performed against selected bacterial strains using the 96-well microplate method with resazurin dye as an indicator. Four clinically relevant and antibiotic-resistant isolates were chosen for testing: Staphylococcus aureus, Pseudomonas aeruginosa, Escherichia coli, and Klebsiella pneumoniae. MIC values were determined to assess the inhibitory effects of the purified compounds, as summarized in Table 1 and Figure 1.
B. Minimum bactericidal concentration (MBC)
The MBC is defined as the lowest concentration of an antimicrobial agent required to kill 99.9% of bacteria, determined after subculturing the treated organism into antibiotic-free media [19] as summarized in Table 2 and Figure 2.
Table 2. MBC of selected bioactive compounds (µg/mL) against uropathogenic isolates
|
Bacterial Isolate |
Oleic Acid |
Linoleic Acid |
Ascorbic Acid |
Dibutyl Phthalate |
Diethyl Phthalate |
Cis-Vaccenic Acid |
P-value |
|
Escherichia coli |
1024 ± 18.7 |
1024 ± 18.7 |
1024 ± 18.7 |
1024 ± 18.7 |
512 ± 12.7 |
256 ± 4.5 |
0.0001 ** |
|
Klebsiella pneumoniae |
1024 ± 18.7 |
512 ± 12.7 |
512 ± 12.7 |
512 ± 12.7 |
512 ± 12.7 |
128 ± 2.9 |
0.0001 ** |
|
Pseudomonas aeruginosa |
512 ± 12.7 |
2048 ± 32.6 |
2048 ± 32.6 |
2048 ± 32.6 |
1024 ± 18.7 |
128 ± 2.9 |
0.0001 ** |
|
Staphylococcus aureus |
512 ± 12.7 |
512 ± 12.7 |
512 ± 12.7 |
1024 ± 18.7 |
256 ± 4.5 |
64 ± 1.04 |
0.0001 ** |
Note: Data as Mean ± SD are expressed (n = 3). ** indicates significant difference at P ≤ 0.01.
Figure 2. MBC values of bioactive compounds against the uropathogenic isolate
3.2 Anticancer activity of purified cis-vaccenic acid and diethyl phthalate produced by Bacillus thermoamylovorans against human MCF-7 and HeLa cell lines
3.2.1 Cytotoxic effect of cis-vaccenic acid from Bacillus thermoamylovorans on MCF-7 and HeLa cells using MTT assay
The cytotoxic activity of purified cis-vaccenic acid and diethyl phthalate, produced from Bacillus thermoamylovorans, was evaluated against MCF-7 and HeLa cancer cell lines using the3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) assay. This endpoint colorimetric assay measures cell viability at specific time points, allowing for the determination of IC₅₀ values. Cell viability was measured at each time point using the MTT assay. Results presented in Tables 3 and 4 indicate that purified diethyl phthalate (1.12 g/mL) exhibited potent cytotoxic activity against both MCF-7 and HeLa cells. A sigmoidal dose–response curve was fitted to the experimental data, and the IC₅₀ value was determined graphically from the fitted curve. As shown in Figures 3-6, the IC₅₀ value of purified diethyl phthalate after 72 h of incubation at 37℃ was 445 μg/mL for MCF-7 cells and 369 μg/mL for HeLa cells.
Table 3. The cytotoxic effect of diethyl phthalate on the MCF-7 cell line
|
Concentration (1.12 g/mL) |
OD (Mean ± SD) |
Cell Viability (%) |
|
1000 |
0.097 ± 0.014 |
15 |
|
500 |
0.254 ± 0.020 |
39 |
|
250 |
0.572 ± 0.048 |
88 |
|
100 |
0.540 ± 0.013 |
83 |
|
50 |
0.546 ± 0.017 |
84 |
|
25 |
0.577 ± 0.043 |
89 |
|
10 |
0.598 ± 0.027 |
92 |
|
5 |
0.648 ± 0.024 |
100 |
|
1 |
0.650 ± 0.011 |
100 |
|
0 (Control) |
0.651 ± 0.017 |
100 |
Values as mean ± standard deviation (SD) are expressed from four independent measurements (n = 4). IC₅₀ values were calculated based on cell viability percentages.
Table 4. The cytotoxic effect of diethyl phthalate on the HeLa cell line
|
Concentration (1.12 g/mL) |
OD (Mean ± SD) |
Cell Viability (%) |
|
1000 |
0.078 ± 0.017 |
20 |
|
500 |
0.082 ± 0.007 |
20 |
|
250 |
0.333 ± 0.029 |
83 |
|
100 |
0.360 ± 0.022 |
90 |
|
50 |
0.365 ± 0.013 |
91 |
|
25 |
0.372 ± 0.026 |
93 |
|
10 |
0.369 ± 0.006 |
92 |
|
5 |
0.379 ± 0.004 |
95 |
|
1 |
0.385 ± 0.028 |
96 |
|
0 (Control) |
0.399 ± 0.025 |
100 |
Values as mean ± standard deviation (SD) are expressed from four independent measurements (n = 4). IC₅₀ values were calculated based on cell viability percentages.
Figure 3. Cytotoxicity effect of diethyl phthalate on the viability of the MCF-7 cell line at different concentrations
Figure 4. Cytotoxic effect of diethyl phthalate on the viability of MCF-7 cell line expressed as Log concentration (µg/mL)
Figure 5. Cytotoxicity effect of diethyl phthalate on the viability of the MCF-7 cell line at different concentrations
Figure 6. Cytotoxic effect of diethyl phthalate on the viability of the HeLa cell line expressed as Log concentration (µg/mL)
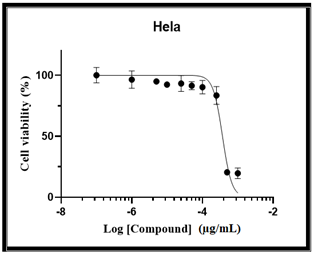
The cytotoxic activity of purified cis-vaccenic acid was also examined against MCF-7 and HeLa cells. As presented in Tables 5 and 6, treatment with 100 mg/mL of purified cis-vaccenic acid showed a strong cytotoxic effect on both cancer cell lines. The IC₅₀ values, determined after 48 h of incubation at 37℃, were 20 μg/mL for MCF-7 cells and 12.9 μg/mL for HeLa cells, as illustrated in Figures 7, 8, and 9.
Table 5. The cytotoxic effect of cis-vaccenic acid on the MCF-7 cell line
|
Concentration (100 mg/mL) |
OD (Mean ± SD) |
Cell Viability (%) |
|
100 |
0.106 ± 0.021 |
18 |
|
50 |
0.110 ± 0.022 |
18 |
|
25 |
0.346 ± 0.032 |
57 |
|
10 |
0.354 ± 0.037 |
58 |
|
5 |
0.558 ± 0.011 |
92 |
|
1 |
0.528 ± 0.007 |
87 |
|
0.5 |
0.542 ± 0.026 |
89 |
|
0 (Control) |
0.607 ± 0.033 |
100 |
Values as mean ± standard deviation (SD) are presented for four independent measurements (n = 4). IC₅₀ was calculated based on cell viability (%).
Table 6. The cytotoxic effect of cis-vaccenic acid on the HeLa cell line
|
Concentration (100 mg/mL) |
OD (Mean ± SD) |
Cell Viability (%) |
|
100 |
0.071 ± 0.003 |
17 |
|
50 |
0.072 ± 0.004 |
17 |
|
25 |
0.068 ± 0.005 |
16 |
|
10 |
0.359 ± 0.029 |
84 |
|
5 |
0.385 ± 0.004 |
91 |
|
1 |
0.359 ± 0.008 |
84 |
|
0.5 |
0.355 ± 0.023 |
84 |
|
0 (Control) |
0.425 ± 0.014 |
100 |
Values as mean ± standard deviation (SD) are presented for four independent measurements (n = 4). IC₅₀ was calculated based on cell viability (%).
Figure 7. Cytotoxic effect of cis-vaccenic acid on the viability of the MCF-7 cell line at different concentrations
Figure 8. Cytotoxic effect of cis-vaccenic acid on the viability of the HeLa cell line at different concentrations
Figure 9. Cytotoxic effect of cis-vaccenic acid on the viability of MCF-7 cell lines expressed as Log concentration (µg/mL)
This study provides the first integrated evaluation of the anticancer and antibacterial activities of bioactive metabolites by Bacillus thermoamylovorans, with particular emphasis on cis-vaccenic acid (CVA) and diethyl phthalate (DEP). Our findings revealed three major thematic insights:
(1) the exceptional potency of cis-vaccenic acid,
(2) the variation in susceptibility among different bacterial species, and
(3) the potential mechanistic overlap between antibacterial and anticancer effects.
Through all assays, cis-vaccenic acid exhibited the strongest antibacterial and anticancer activities. The superior performance of CVA is linked plausibly of unsaturated fatty acids to the biological characteristics, their ability particularly to integrate into and destabilize lipid bilayers. Many studies illustrated that monounsaturated fatty acids, involving CVA and oleic acid, increase permeability, disrupt membrane integrity, and induce leakage of intracellular contents in both cancer cells and bacteria [20, 21]. In organisms lacking robust membrane-reinforcing mechanisms, such effects are amplified. Furthermore, intracellular redox homeostasis may be modulated across CVA. The recent reports show that unsaturated fatty acids can increase the formation of reactive oxygen species (ROS) and disrupt mitochondrial function, leading to in various cancer cells apoptotic cell death pathways [22]. These mechanisms observed align with the markedly decreased IC₅₀ values in HeLa and MCF-7 cell lines, which were substantially lower than those of DEP. The increased sensitivity of HeLa cells may be due to a higher dependency on mitochondrial metabolism, making them more vulnerable to ROS-mediated apoptosis. Consistent with these mechanisms, cis-vaccenic acid exhibited a clear dose-dependent reduction in cell viability, with significant cytotoxic effects observed at concentrations ≥ 25 µg/mL.
Additionally, the antibacterial effect of CVA is strong against Klebsiella pneumoniae and Staphylococcus aureus supports evidence that fatty acids penetrate more easily into envelopes of Gram-positive cells, due to structural collapse of the cytoplasmic membrane [20]. Overall, CVA with substantial antimicrobial and cytotoxic potential emerges as a dual-functional metabolite. The tested uropathogens displayed significant variation in their susceptibility significant variation to the evaluated compounds. These differences are attributable largely to physiological and structural characteristics. Gram-negative bacteria like Pseudomonas aeruginosa and E. coli possess a rich outer membrane in lipopolysaccharides and form a robust permeability barrier, efflux pumps against hydrophobic molecules, involving phthalate derivatives and fatty acids. The complexity of the structure explains why their sensitivity is reduced and requires high concentrations for inhibition, with previous reports consistent [23].
In contrast, Gram-positive bacteria, S. aureus, particularly exhibited markedly higher sensitivity to CVA and DEP. An outer membrane absence and a more permeable peptidoglycan layer presence facilitate the entry of lipid-active compounds. The susceptibility of S. aureus to CVA (lowest MIC and MBC values) suggests a mechanism including disruption of membrane potential rapidly and ion homeostasis, for related fatty acids as described [20]. Interestingly, P. aeruginosa, with its high intrinsic resistance to other compounds, remained sensitive to CVA relative. This may reflect CVA's ability to overcome outer-membrane resistance, bypass or interfere with membrane-bound enzymes essential for survival. To vaccenic acids, similar strain-dependent sensitivity has recently been documented [10].
A notable outcome of this study of CVA is the parallel pattern potency in both antibacterial and anticancer. Although speculative, overlapping biological targets may reflect this correlation. Both bacteria and cancer cells affect membrane integrity, energy of metabolism, and redox balance. Compounds that induce oxidative stress and disrupt membranes in bacteria may exert similar effects in cancer cells, as both systems are susceptible to lipid peroxidation and mitochondrial dysfunction are susceptible [24]. Indeed, ROS apoptotic and generation signaling by CVA are shared pathways activated, at high concentrations, by DEP. Overall, the consistency across microbial and cancer models in biological activity suggests that metabolites, particularly CVA from B. thermoamylovorans, for developing multifunctional therapeutic agents may serve as promising scaffolds.
This study focuses on the therapeutic potential of bioactive secondary metabolites produced by Bacillus thermoamylovorans. Among the compounds evaluated, cis-vaccenic acid was found to have the most potent antibacterial and anticancer activities exhibited, likely due to its ability to influence oxidative and disrupt membrane structures and mitochondrial pathways. Variations in the sensitivity of bacteria were associated strongly with differences in resistance mechanisms and cell-wall architecture. In both antibacterial and anticancer models, the parallel efficacy of CVA suggests possible mechanistic overlap, although this needs through targeted biochemical assays further validation. Collectively, as a promising source of biologically active metabolites, the findings position B. thermoamylovorans to explore their therapeutic relevance justify deeper molecular investigations.
The special thanks from the authors to Mustansiriyah University, College of Science, and Department of Biology, Baghdad, Iraq.
[1] Beladjal, L., Gheysens, T., Clegg, J.S., Amar, M., Mertens, J. (2018). Life from the ashes: Survival of dry bacterial spores after very high temperature exposure. Extremophiles, 22(5): 751-759. https://doi.org/10.1007/s00792-018-1035-6
[2] Errington, J., Aart, L.T.V.D. (2020). Microbe Profile: Bacillus subtilis: Model organism for cellular development, and industrial workhorse. Microbiology, 166(5): 425-427. https://doi.org/10.1099/mic.0.000922
[3] Krawczyk, A.O., Berendsen, E.M., Eijlander, R.T., de Jong, A., Wells-Bennik, M.H., Kuipers, O.P. (2015). Draft genome sequences of four Bacillus thermoamylovorans strains isolated from milk and acacia gum, a food ingredient. Genome Announcements, 3(2): e00165-15. https://doi.org/10.1128/genomea.00165-15
[4] Nigris, S., Baldan, E., Tondello, A., Zanella, F., et al. (2018). Biocontrol traits of Bacillus licheniformis GL174, a culturable endophyte of Vitis vinifera cv. Glera. BMC Microbiology, 18(1): 133. https://doi.org/10.1186/s12866-018-1306-5
[5] Sumi, C.D., Yang, B.W., Yeo, I.C., Hahm, Y.T. (2015). Antimicrobial peptides of the genus Bacillus: A new era for antibiotics. Canadian Journal of Microbiology, 61(2): 93-103. https://doi.org/10.1139/cjm-2014-0613
[6] Aqel, H., Farah, H., Al-Hunaiti, A. (2024). Ecological versatility and biotechnological promise: Comprehensive characterization of the isolated thermophilic Bacillus strains. Plos One, 19(4): e0297217. https://doi.org/10.1371/journal.pone.0297217
[7] Gerbec, Z.J., Serapio-Palacios, A., Metcalfe-Roach, A., Krekhno, Z., et al. (2025). Identification of intratumoral bacteria that enhance breast tumor metastasis. mBio, 16(3): e03595-24. https://doi.org/10.1128/mbio.03595-24
[8] Mohammad, M.K., Hussain, S.S., Mahdi, A.S. (2025). GC-MS characterization of antibacterial compounds from Bacillus thermoamylovorans isolated from human skin microbiome. International Journal of Design & Nature and Ecodynamics, 20(9): 2027-2036. https://doi.org/10.18280/ijdne.200908
[9] Piwowarek, K., Lipińska, E., Kieliszek, M. (2023). Reprocessing of side-streams towards obtaining valuable bacterial metabolites. Applied Microbiology and Biotechnology, 107(7): 2169-2208. https://doi.org/10.1007/s00253-023-12458-8
[10] Maumela, P., Serepa-Dlamini, M.H. (2024). In silico analysis of novel bacterial metabolites with anticancer activities. Metabolites, 14(3): 163. https://doi.org/10.3390/metabo14030163
[11] Flores-Mireles, A.L., Walker, J.N., Caparon, M., Hultgren, S.J. (2015). Urinary tract infections: Epidemiology, mechanisms of infection and treatment options. Nature Reviews Microbiology, 13(5): 269-284. https://doi.org/10.1038/nrmicro3432
[12] Pichler, M., Steyrer, J. (2021). Cost-effectiveness analysis of the use of immunotherapy in metastatic solid tumours in Austria by applying the ESMO-Magnitude of Clinical Benefit Scale (ESMO-MCBS) version 1.1. ESMO Open, 6(4): 100198. https://doi.org/10.1016/j.esmoop.2021.100198
[13] Capes-Davis, A., Theodosopoulos, G., Atkin, I., Drexler, H.G., et al. (2010). Check your cultures! A list of cross‐contaminated or misidentified cell lines. International Journal of Cancer, 127(1): 1-8. https://doi.org/10.1002/ijc.25242
[14] Yazıcı, A. (2024). The strain-dependent antimicrobial and antibiofilm effect of cis and trans-vaccenic acid against Pseudomonas aeruginosa. Cumhuriyet Science Journal, 45(1): 1-7.
[15] Ozyigit, I.I., Dogan, I., Hocaoglu-Ozyigit, A., Yalcin, B., Erdogan, A., Yalcin, I.E., Cabi, E., Kaya, Y. (2023). Production of secondary metabolites using tissue culture-based biotechnological applications. Frontiers in Plant Science, 14: 1132555. https://doi.org/10.3389/fpls.2023.1132555
[16] Massadeh, M.I., Al Masri, M. (2022). Bioactive secondary metabolites production by terrestrial Bacillus circulans strains. Journal of Microbiology, Biotechnology and Food Sciences, 12(2): e5666. https://doi.org/10.55251/jmbfs.5666
[17] Tan, B., Zhao, H., Du, L., Gan, X., Quan, X. (2016). A versatile fluorescent biosensor based on target-responsive graphene oxide hydrogel for antibiotic detection. Biosensors and Bioelectronics, 83: 267-273. https://doi.org/10.1016/j.bios.2016.04.065
[18] Gummlich, L., Kaehne, T., Naumann, M., Kilic, E., Jung, K., Dubiel, W. (2016). New insights into the mechanism of COP9 Signalosome–Cullin-RING ubiquitin-ligase pathway deregulation in urological cancers. International Review of Cell and Molecular Biology, 323: 181-229. https://doi.org/10.1016/bs.ircmb.2015.12.007
[19] Ohikhena, F.U., Wintola, O.A., Afolayan, A.J. (2017). Evaluation of the antibacterial and antifungal properties of Phragmanthera capitata (Sprengel) Balle (Loranthaceae), a mistletoe growing on rubber tree, using the dilution techniques. The Scientific World Journal, 2017(1): 9658598. https://doi.org/10.1155/2017/9658598
[20] Yoon, B.K., Jackman, J.A., Valle-González, E.R., Cho, N.J. (2018). Antibacterial free fatty acids and monoglycerides: Biological activities, experimental testing, and therapeutic applications. International Journal of Molecular Sciences, 19(4): 1114. https://doi.org/10.3390/ijms19041114
[21] Xu, R., Molenaar, A.J., Chen, Z., Yuan, Y. (2025). Mode and mechanism of action of Omega-3 and Omega-6 unsaturated fatty acids in chronic diseases. Nutrients, 17(9): 1540. https://doi.org/10.3390/nu17091540
[22] Song, J., Wang, Y., Fan, X., Wu, H., Han, J., Yang, M., Lu, L., Nie, G. (2019). Trans-vaccenic acid inhibits proliferation and induces apoptosis of human nasopharyngeal carcinoma cells via a mitochondrial-mediated apoptosis pathway. Lipids in Health and Disease, 18(1): 46. https://doi.org/10.1186/s12944-019-0993-8
[23] Subramaniam, R., Rahma, S.B.A., Taib, W.R.B.W., Zahri, M.K. (2021). The cytotoxicity effect on MCF-7 human breast cancer cell line treated with Prismatomeris glabra leaves and roots ethyl acetate extract. Asian Journal of Medicine and Biomedicine, 5(S1): 47-53. https://doi.org/10.37231/ajmb.2021.5.S1.448
[24] Casillas-Vargas, G., Ocasio-Malavé, C., Medina, S., Morales-Guzmán, C., Del Valle, R.G., Carballeira, N.M., Sanabria-Ríos, D.J. (2021). Antibacterial fatty acids: An update of possible mechanisms of action and implications in the development of the next-generation of antibacterial agents. Progress in Lipid Research, 82: 101093. https://doi.org/10.1016/j.plipres.2021.101093