Ni Wayan Febriana Utami*![]() | Gede Mekse Korri Arisena
| Gede Mekse Korri Arisena![]() | Ni Luh Made Indah Murdyani Dewi
| Ni Luh Made Indah Murdyani Dewi![]() | Ni Wayan Sri Waras Danu Dewi
| Ni Wayan Sri Waras Danu Dewi![]() | Ary Bakhtiar
| Ary Bakhtiar![]() | Pande Made Ari Ananta Paramarta
| Pande Made Ari Ananta Paramarta![]()
© 2025 The authors. This article is published by IIETA and is licensed under the CC BY 4.0 license (http://creativecommons.org/licenses/by/4.0/).
OPEN ACCESS
The Tukad Ayung Watershed, the longest in Bali Province, functions as a vital buffer zone for food production areas. However, variation in vegetation composition across its upstream, midstream, and downstream regions has not been adequately examined in relation to ecosystem service capacity. This study aims to (1) analyze tree vegetation diversity, (2) assess its variation based on species origin (native/exotic) and growth status (planted/naturally grown), and (3) analyze aboveground tree biomass contribution to support sustainable food production. Vegetation diversity was evaluated using the Shannon–Wiener index, and basal area calculations were used to estimate aboveground tree biomass and carbon sequestration. Statistical significance in the distribution of tree origin, tree status, and biomass among the three villages (Belancan, Kerta, and Abiansemal) was tested using Pearson’s Chi-square test, while data visualization was performed with the ggplot2 package in R software version 4.3.3. The results showed Shannon–Wiener diversity indices (H') of 3.01 in Belancan Village, 2.79 in Abiansemal Village, and 2.67 in Kerta Village. Vegetation composition varied spatially, with endemic dominance in Belancan and Abiansemal and exotic dominance in Kerta driven by human cultivation. Aboveground tree biomass was highest in Abiansemal (704.81 kg/ha) and lowest in Kerta (380.37 kg/ha). The chi-square test indicated significant differences in biomass (χ² = 582.76, df = 538, p = 0.089) among locations. These findings demonstrate that the Tukad Ayung Watershed has high vegetation diversity and biomass potential, emphasizing the importance of site-specific conservation and mixed-garden management strategies to maintain ecosystem stability and food security.
biomass, biodiversity, food security, Tukad Ayung, watershed management
Agriculture plays a crucial role in supporting global food security and rural livelihoods. However, increasing urbanization and land-use change have transformed many agrarian landscapes into built-up areas, leading to the degradation of natural ecosystems and the loss of biodiversity and genetic resources [1, 2]. In tropical regions such as Indonesia, these transformations frequently occur within watershed systems, where agricultural landscapes are replaced or fragmented by settlements and infrastructure development [3]. Such changes not only reduce agricultural productivity but also disrupt ecological processes essential for maintaining watershed functions.
A watershed consists of interconnected upstream, midstream, and downstream zones that regulate hydrological and ecological processes [4, 5]. Land-use change within these zones alters sediment dynamics, vegetation structure, and habitat integrity, leading to soil erosion, sedimentation, and reduced ecosystem services [6, 7]. While these biophysical impacts of urbanization are well-documented [8], less attention has been paid to how changes in vegetation composition [9], particularly within agroforestry systems such as mixed gardens which affect biodiversity and ecological function at the watershed scale.
In Bali, rapid urban growth and tourism-driven land conversion have significantly altered traditional agricultural landscapes, including those within the Tukad Ayung Watershed. This watershed, which spans the regencies of Bangli, Gianyar, Tabanan, and Denpasar City, covers approximately 30,981 hectares and supports diverse land uses including riparian zones, mixed gardens, and forest patches, all of which play a crucial role in maintaining ecosystem stability [8]. Increasing land conversion and monoculture practices have resulted in biodiversity loss, declining carbon storage, and reduced erosion control capacity and other ecosystem services [3].
Abian mixed-gardens, a form of traditional dry farmland agroforestry in Bali [10, 11], play an essential ecological role by maintaining vegetation diversity, providing biomass, and supporting carbon sequestration. However, the relationship between vegetation diversity, tree characteristics, and their ecological functions across watershed zones remains insufficiently understood. Addressing this gap is critical for developing effective conservation and land management strategies that balance ecological sustainability with food production, which is the novelty of this study.
Therefore, this research investigates vegetation diversity and ecological functions across upstream, midstream, and downstream zones of the Tukad Ayung Watershed. Specifically, it aims to (i) analyze vegetation diversity across watershed zones, (ii) determine if there are significant differences in the proportion of tree origin (endemic/exotic) and growth status (planted/naturally grown) across the watershed zones, and (iii) assess aboveground tree biomass and carbon storage contribution to ecosystem services. These findings are expected to inform locally adapted conservation strategies and sustainable mixed garden management to maintain the ecological integrity of the Tukad Ayung Watershed.
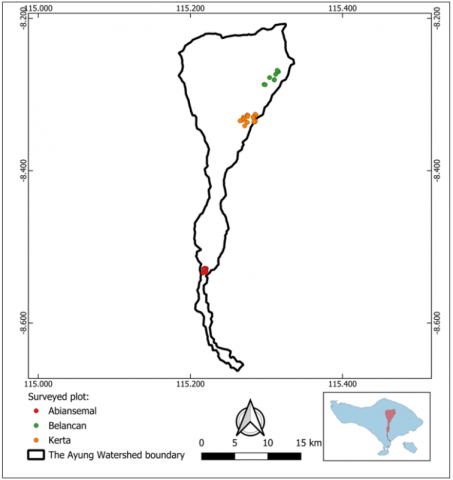
This study employed a field survey method to estimate tree species diversity, differences, and aboveground tree biomass and carbon storage contribution in the three watershed zones. The selection of sample villages was carried out using purposive sampling, based on representativeness and regional accessibility. In each watershed zone, 20 mixed gardens were surveyed, with one 10 × 10 m sample plot established in each garden. A total of 60 sample plots were therefore established across the three watershed zones: upstream, midstream, and downstream (Figure 1).
The study design, as described was provided wide spatial coverage and allowed for comparative analysis of vegetation diversity and aboveground tree biomass and carbon storage across watershed gradients, following standard approaches in tropical agroforestry studies [12, 13].
In each sample plot, a tree census was conducted by recording species name, tree height, and diameter at breast height (DBH) for trees with DBH > 5 cm [14]. Additional information recorded included plant status (planted/naturally grown) and plant origin (native or exotic) to reflect the human planting and management [15, 16]. Data analysis was divided into three parts. First, the analysis of tree stratum vegetation diversity was assessed using the Shannon–Wiener index [17-19], calculated as follows:
$\mathrm{H}^{\prime}=-\sum_{i=1}^S(P i \operatorname{Ln} P i)$ (1)
where,
H’: Species diversity index;
S: Total number of species;
Pi: Proportion of individuals of species i to the total number of individuals;
Ln: Natural logarithm in i.
Based on the Shannon–Wiener diversity index, values can be categorized as follows: H’< 1 indicates low diversity; 1< H’< 3 indicates moderate diversity; and H’>3 indicates high diversity.
The second objective was to determine if there are significant differences in the proportion of tree origin (endemic/exotic) and growth status (planted/naturally grown) [20]. This objective was analyzed using Pearson’s Chi-squared test, calculated with the following formula:
$\chi^2=\sum \frac{\left(O_i-E_i\right)^2}{E_i}$ (2)
where,
χ² = chi-squared;
Oi = observed value;
Ei = expected value.
The third objective was to assess the contribution of above-ground tree biomass and carbon sequestration across watershed zones. Basal area (BA) was first calculated to describe stand structure and species dominance within each mixed garden plot, but was not directly used for aboveground tree biomass estimation. Aboveground tree biomass (W) was estimated directly from DBH using the generalized allometric model developed by Sandoya et al. [20], for tropical forests and agroforestry systems, as species-specific equations were unavailable. The carbon content (C) was estimated as 50% of aboveground tree biomass, following the default carbon fraction commonly used for tropical vegetation [20, 21].
$W=a D^b$ (3)
$\mathrm{C}=0.5 \mathrm{~W}$ (4)
where,
W: Aboveground tree biomass (kg/ha);
C: Carbon content (kg/ha);
a, b: 0.19 and 2.37, respectively (constant derived from Brown [21]).
Following data analysis, visualization of the aboveground species biomass in the upstream, midstream, and downstream zones of the Tukad Ayung Watershed was performed using the ggplot2 package [22] in R software (version 4.3.3) [23]. Statistical significance was assessed using Pearson’s Chi-squared test, and the results were visualized in the same environment for cross-location comparisons [24]. In addition, the chi-square (χ²) test was applied to examine whether the distribution of tree species based on origin (native or exotic) and status (planted or naturally grown) categories differed significantly among the three villages (Belancan, Kerta, and Abiansemal). All statistical a were conducted in R software (version 4.3.3) with significance thresholds set at p < 0.05.
3.1 Vegetation diversity status across the Tukad Ayung Watershed
Vegetation diversity reflects the variation in plant species composition within a particular area or ecosystem. Based on the Shannon–Wiener diversity index (H’) of trees in the Tukad Ayung Watershed, Belancan Village recorded the highest biodiversity value at 3.01, followed by Abiansemal Village at 2.79, and Kerta Village at 2.67, all of which are categorized as highly diverse (Table 1). High diversity index values are generally found in areas with minimal human intervention, such as upstream zones adjacent to forested regions [25]. The relatively high H’ values in the upstream, midstream, and downstream zones indicate a fairly good diversity of tree species, although the degree of disturbance varies among zones. The persistence of high diversity despite human influence suggests that traditional Abian mixed-garden practices play an important role in maintaining vegetation heterogeneity and ecosystem stability [11]. Therefore, conservation efforts in the Tukad Ayung Watershed should prioritize strategies that consider the characteristics of the local agroforestry management system and its role in maintaining ecological stability.
Table 1. Comparison of biodiversity index (H’) and diversity status in the Tukad Ayung Watershed
|
Watershed Zones (Representative Village) |
Diversity Index (H’) |
Diversity Status |
|
Upstream (Belancan)*** |
3.01 |
highly diverse |
|
Midstream (Kerta)* |
2.67 |
highly diverse |
|
Downstream (Abiansemal)** |
2.79 |
highly diverse |
Description: The star symbol indicates the diversity value ranking among three locations. Belancan (***) has a slightly higher value than Abiansemal (**), and Kerta (*) has the lowest among the other two locations.
From Table 1, it is evident that Belancan Village, located in the upstream area of the Tukad Ayung Watershed, exhibits a highly diverse index with the highest H’ value of 3.01 among the other two locations. Tree vegetation diversity in Belancan Village comprises 37 tree species, dominated by avocado (Persea americana, 5.1%), tangerine (Citrus nobilis, 4.1%), and jackfruit (Artocarpus heterophyllus, 3%). Avocado was planted because it relies on plantations that have economically valuable crops to support their livelihood. Avocado is widely cultivated in all plots in the upstream area because it has a high market demand [26]. Another tree species found is the jackfruit. Villagers in Belancan choose to plant A. heterophyllus because it has a cultural significance in Balinese society, being widely used in traditional ceremonies and daily life [27], aside from its economic value.
Mixed gardens in Kerta Village consist of 27 tree species and have an H’ value of 2.67, which is still categorized as highly diverse (Table 1), even though it is the lowest among the three locations. The vegetation found in the Kerta mixed garden sample plot is dominated by tangerine (Citrus nobilis), which accounts for 12% of the trees. Tangerine is a major fruit commodity with considerable global production and trade [28]. The dominance of C. nobilis plantation reflects more intensive land-use management for commercial production, particularly of fruit crops. These trends indicate that market-oriented cultivation, as observed in Kerta Village, has increased human influence on vegetation composition and reduced naturally regenerating tree species.
The lowest watershed village sampled, Abiansemal, has an H’ index value of 2.79 and is categorized as highly diverse (Table 1). The Abian mixed garden sample plots in Abiansemal contain 30 tree species, with coconut (Cocos nucifera) comprising the highest proportion of individuals at 23%. The high number of coconut trees observed is similar to the pattern for jackfruit, as both species hold deep cultural importance in Balinese society and are essential components of religious and daily ceremonies [27]. This finding aligns with observations from other tropical regions, such as India, where C. nucifera also carries strong cultural significance in local culinary, health practices, and spiritual life [29]. Therefore, coconut not only contributes to ecological balance and biodiversity enhancement but also represents a vital link between environmental and cultural sustainability.
3.2 Identification of significant differences based on tree origin (endemic/exotic) and growth status (cultivated/naturally grown)
The theory underlying differences and composition of vegetation in mixed gardens in the upstream, midstream, and downstream areas of the Tukad Ayung Watershed is closely related to the concept of landscape ecology and ecosystem stratification within watershed regions. Within watershed systems, the upstream, midstream, and downstream areas possess distinct biophysical and socio-economic characteristics, which in turn influence the selection and presence of vegetation types [30, 31]. The variation in vegetation composition across the upstream, midstream, and downstream areas of the Tukad Ayung Watershed is the result of interactions between biophysical conditions and socio-economic factors, thereby necessitating integrated consideration of both aspects in ecosystem management [32, 33].
The characteristics of plants in the Tukad Ayung Watershed are viewed in terms of tree origin and status, reflecting the diversity of provenance as well as ecological roles within mixed gardens. Endemic plants are native species found only in specific regions, playing a crucial role in maintaining the balance of local ecosystems [34]. Conversely, exotic plants are introduced from outside regions or countries, generally due to economic or aesthetic value, but may become invasive if not properly managed. Cultivated plants are deliberately grown by communities, typically for economic, cultural, or food purposes [35]. Meanwhile, naturally growing plants emerge through natural regeneration, demonstrating resilience to local environmental conditions [34]. The distinction between exotic, cultivated, and naturally growing plants highlights that economic, cultural, and ecological adaptation factors are central in shaping vegetation composition, thereby underscoring the need for ecosystem sustainability management to take these three aspects into account.
3.2.1 Tree origin in mixed gardens
Differences and compositions of vegetation in mixed gardens display specific ecological characteristics, namely, environmental conditions and resources that support survival, growth, and reproduction. In upstream areas, which are generally characterized by steep topography, high rainfall, and relatively natural soils, species adapted to such conditions, such as local trees with strong root systems and resilience in nutrient-poor soils, tend to dominate [36]. More moderate environmental conditions in midstream areas allow for higher interspecies competition, leading to optimal diversity in mixed gardens. In downstream areas, greater anthropogenic pressures and more open land result in dominance by species that are competitive in utilizing light and space, such as economically valuable fruit trees.
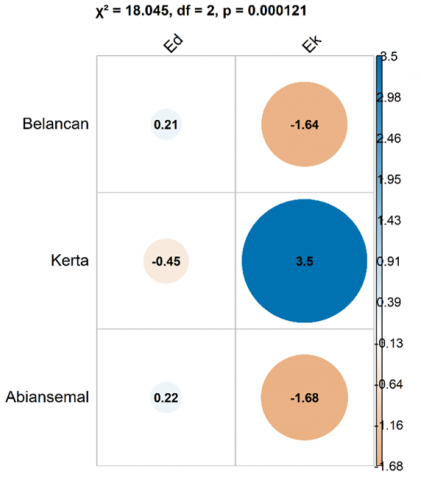
The residual Chi-square results, which measure each area’s contribution to the distribution of species types (endemic and exotic), show that Belancan and Abiansemal Villages exhibit a higher dominance of endemic (Ed) species (0.21 and 0.2, respectively), whereas Kerta Village demonstrates a concentration of exotic (Ek) species (3.50), indicating a significantly higher presence than expected (Figure 2). This interpretation reinforces the understanding that each region has distinct ecological characteristics and contributes differently to the distribution of endemic (Ed) and exotic (Ek) species, which is crucial for decision-making in conservation and local ecosystem management [37]. The dominance of endemic species in Belancan and Abiansemal Villages, along with the concentration of exotic species in Kerta Village, highlights ecological differences among regions, suggesting that conservation strategies and ecosystem management must be tailored to the specific contributions of each village to species distribution.
Figure 2. Corrplot of tree origin in the upstream, midstream, and downstream areas of the Tukad Ayung Watershed
The Chi-square test results indicate a significant difference in the distribution of endemic tree origin status among the three regions of the Tukad Ayung Watershed—upstream, midstream, and downstream (Pearson’s Chi-squared = 18.045; df = 2; p-value < 0.000121), suggesting variation in vegetation composition across regions [36]. The composition of mixed garden vegetation is strongly influenced by geographical factors (e.g., altitude and climate), cultural aspects (local preferences), as well as the economic needs of local communities [38]. Understanding these differences is crucial for designing conservation and sustainable agricultural strategies that align with the local characteristics of each watershed region [20]. The statistical significance demonstrated across the three locations (p-value = 2.2e-16) indicates that environmental, anthropogenic, or specific ecological conditions of each region are not incidental, but rather shaped by the stability of local ecosystems [36]. This finding warrants special attention in the management of biodiversity and its implications for the conservation of mixed garden ecosystems (Abian) in Bali [3]. The significant differences in vegetation composition across the upstream, midstream, and downstream areas of the Tukad Ayung Watershed reflect the influence of geographical, cultural, and economic factors that must be considered in conservation planning and sustainable management of mixed garden ecosystems.
3.2.2 Tree status in mixed gardens
The characteristics of cultivated trees and naturally grown trees represent two distinct approaches to vegetation management and the utilization of natural resources [39]. Cultivated trees (Ta) are generally selected and managed by humans for specific economic, aesthetic, or functional purposes, such as fruit production, timber, or protective roles [40]. This type often reflects the dominance of particular species aligned with market demands or local cultural preferences. Cultivated trees within agricultural and plantation contexts exemplify planned and controlled cultivation systems, which tend to be more homogeneous. In contrast, naturally grown trees (Tu) arise from natural regeneration processes such as seed dispersal by wind, animals, or other ecological mechanisms [41, 42]. These trees exhibit high adaptability to local environmental conditions and typically display greater species, structural, and functional diversity within the ecosystem. The presence of naturally grown trees, in ecological theory, reflects ecosystem stability and resilience, as they support nutrient cycling, soil moisture, and fauna habitats [38]. Both groups complement each other in mixed gardens, with cultivated trees providing direct economic benefits, while naturally grown trees safeguard ecological functions and long-term sustainability.
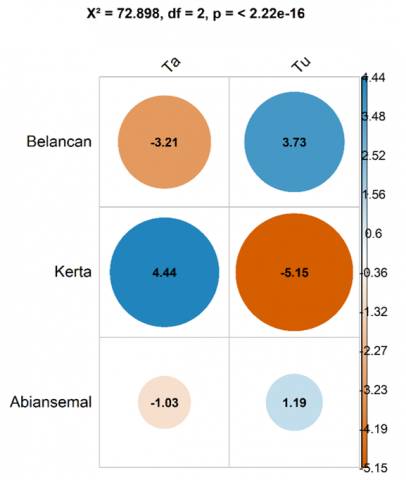
The residuals Chi-square test results show a significant difference in the distribution of cultivated trees and naturally grown trees among the three regions of the Tukad Ayung Watershed, namely upstream, midstream, and downstream (Pearson’s Chi-squared = 72.898; df = 2; p-value < 2.22e-16). This composition may be influenced by the economic needs of local communities in utilizing mixed gardens as income sources [43]. These differences must be thoroughly understood by relevant stakeholders in land conservation efforts. The significant values observed across the three watershed regions (p-value < 2.2e-16) indicate that the conditions within each region are not random but are shaped by ecosystem dynamics, requiring specific attention in conserving naturally growing plants that serve as buffers for the Tukad Ayung Watershed in maintaining ecosystem stability.
The residual Chi-square results, which measure the contribution of upstream, midstream, and downstream areas to tree status (cultivated versus naturally grown), show that Belancan Village contributes strongly to both cultivated (Ta) and naturally grown (Tu) trees. Kerta Village demonstrates a higher contribution to cultivated trees compared to naturally grown ones (4.44), whereas in Abiansemal Village, mixed gardens are dominated by naturally grown trees (1.19) (Figure 3). These values suggest that highland communities utilize mixed gardens as a source of income by planting economically valuable trees. In contrast, downstream areas rely on other food crops as their main economic source [43]. Such differences warrant attention, as they are crucial for formulating policies aimed at environmentally friendly conservation of mixed gardens.
Figure 3. Corrplot of tree status in the upstream, midstream, and downstream areas of the Tukad Ayung Watershed
3.3 Contribution of aboveground tree biomass and carbon sequestration in mixed gardens to support sustainable food production
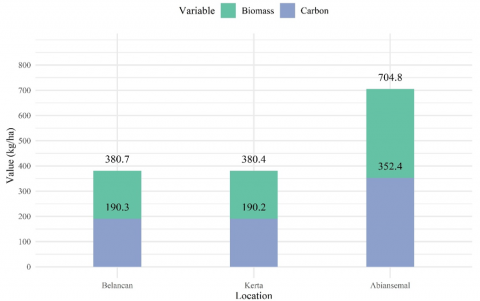
Mixed gardens as an agroforestry system provide various ecosystem services that can be quantified through above-and below-ground biomass, as exemplified by the Abian mixed gardens in Bali [8]. Among the three studied villages, Abiansemal Village recorded the highest aboveground tree biomass in the Tukad Ayung Watershed, amounting to 704.81 kg/ha, far exceeding that of Belancan Village (380.67 kg/ha) and Kerta Village (380.37 kg/ha) (Figure 4). The chi-square (χ²) test for biomass distribution among the villages (χ² = 582.76, df = 538, p = 0.089) indicated no statistically significant difference, suggesting that although mean biomass values varied numerically, these differences were not significant at the 0.05 level. This finding does not necessarily indicate higher human disturbance, as might be expected. Rather, mixed gardens in Abiansemal Village are characterized by denser vegetation and a greater proportion of large, mature trees, which contribute to its higher biomass accumulation.
The greater the total measured biomass, the larger the ecosystem services provided, including carbon storage, biodiversity maintenance, and nutrient cycling, all of which support food security and watershed sustainability. Tree biomass plays a vital ecological, social, and economic role in the Tukad Ayung Watershed, particularly in regions that depend on traditional irrigation systems (Subak) [8]. Tree functions not only as land cover but also as buffers for agricultural sustainability and as a key provider of ecosystem services.
Trees growing around agricultural areas, riverbanks, protected forests, and Abian farmlands generate biomass in the form of trunks, branches, leaves, and roots, which store carbon and enrich the soil with organic matter [2]. Food crops can coexist with timber, fruit, or fodder trees, creating a farming system more resilient to climate change [30]. Moreover, tree biomass provides shade, regulates microclimate humidity, and offers protection from strong winds and heavy rains that can damage crops.
Estimates indicate that carbon sequestration in the Tukad Ayung Watershed reaches 190.3 kg/ha, 190.2 kg/ha, and 352.4 kg/ha in Belancan, Kerta, and Abiansemal, respectively (Figure 4), reflecting the crucial role of vegetation, particularly trees and perennial plants, in absorbing atmospheric CO₂. This capacity demonstrates the watershed’s significant potential to contribute to climate change mitigation through carbon storage in plant tissues [31]. The high carbon sequestration observed in tree species indicates well-maintained vegetation cover and substantial ecosystem service value. A higher rate of carbon sequestration enhances the watershed’s contribution to local climate stabilization, emission reduction, and air quality maintenance.
Figure 4. Tree biomass and carbon storage in the Tukad Ayung Watershed
In the upstream areas, trees growing along slopes and riverbanks help slow rainwater flow before it enters the river, reducing erosion and sedimentation [7]. This function represents a vital ecosystem service that maintains both the quality and quantity of water available to downstream agricultural lands. Trees also assist in filtering groundwater, stabilizing slopes, and absorbing excess nutrients that could pollute rivers [32].
In addition to ecological benefits, tree biomass provides alternative energy sources, such as firewood and charcoal from pruning residues, without compromising the main stands. Products such as fruits, medicinal leaves, and timber enhance farmers’ income and strengthen the economic resilience of communities surrounding the watershed [31]. The long-term integration of trees within the agricultural landscape of the Tukad Ayung Watershed establishes a system more resilient to drought, soil degradation, and flooding. This integration reinforces the role of mixed gardens as a cornerstone of sustainable food production and watershed resilience.
The vegetation diversity of the Tukad Ayung Watershed, based on the Shannon-Wiener index (H'), indicates a high level of diversity, with Belancan Village (upstream) showing the highest values among the three observed locations. This reflects the effective preservation of ecological functions in the watershed, such as sediment control, hydrological balance, and support for sustainable food security among local communities. It also illustrates how local land-use practices, particularly mixed gardens, maintain landscape heterogeneity and ecological resilience.
Tree biomass and estimated carbon storage are highest in Abiansemal Village, reflecting the ecological benefits of maintaining heterogeneous mixed gardens with large, mature trees. Meanwhile, Belancan and Kerta Villages show lower biomass due to the dominance of monocultural or commercially driven planting patterns.
Vegetation composition across the watershed shows spatial variation in tree origin and growth status. Belancan and Abiansemal Villages are dominated by endemic species, while Kerta Village exhibits greater influence from exotic species. Although exotic trees exert ecological pressure in certain zones, overall conservation within the watershed remains effective, with human cultivation patterns—particularly in Kerta—playing a major role in shaping vegetation structure.
This research highlights that mixed gardens function as a critical agroforestry model for integrating biodiversity conservation with productive land use. The spatial variation in tree composition and biomass provides evidence for differentiated management strategies. The upstream region should focus on preserving native vegetation and preventing conversion to intensive cultivation; the midstream area should strengthen the reintroduction of native species to reduce dominance of exotics; and the downstream region should prioritize protecting large, mature trees and improving natural regeneration to restore ecological balance. Therefore, management strategies that locally adapt conservation policy that support sustainable land-use practices and preserve the ecological integrity of the watershed landscape.
We would like to express our gratitude for the research funding provided by the Research Implementation Assignment Agreement (SP3) PNBP fund of Universitas Udayana for the Fiscal Year 2025, Number: B/229.262/UN14.4.A/PT.01.03/2025, dated April 28, 2025 and Budget Implementation List (DIPA) of Universitas Udayana Number: SP DIPA-139.03.2.693389/2025, dated December 2, 2024.
[1] Fanuel, I.M., Mirau, S., Kajunguri, D., Moyo, F. (2023). Conservation of forest biomass and forest–dependent wildlife population: Uncertainty quantification of the model parameters. Heliyon, 9(6): e16948. https://doi.org/10.1016/j.heliyon.2023.e16948
[2] Raihan, A., Begum, R.A., Mohd Said, M.N., Pereira, J.J. (2021). Assessment of carbon stock in forest biomass and emission reduction potential in Malaysia. Forests, 12(10): 1294. https://doi.org/10.3390/f12101294
[3] Eryani, I.G.A.P., Nurhamidah, N. (2020). Sedimentation management strategy in river estuary for control the water damage in downstream of Ayung River. International Journal on Advanced Science Engineering Information Technology, 10(2): 743-748. https://doi.org/10.18517/ijaseit.10.2.10832
[4] Sidek, L.M., Mohiyaden, H.A., Marufuzzaman, M., Noh, N.S.M., Heddam, S., Ehteram, M., Kisi, O., Sammen, S.S. (2024). Developing an ensembled machine learning model for predicting water quality index in Johor River Basin. Environmental Sciences Europe, 36(1): 67. https://doi.org/10.1186/s12302-024-00897-7
[5] Lamsal, A., Germain, R.H., Bevilacqua, E. (2024). Socioeconomic implications of upstream–downstream relationships and watershed communities: A New York City watershed case study. Water, 16(22): 3197. https://doi.org/10.3390/w16223197
[6] Edmonds, D.A., Toby, S.C., Siverd, C.G., Twilley, R., Bentley, S.J., Hagen, S., Xu, K. (2023). Land loss due to human-altered sediment budget in the Mississippi River Delta. Nature Sustainability, 6(6): 644-651. https://doi.org/10.1038/s41893-023-01081-0
[7] Kalsido, T., Berhanu, B. (2020). Impact of land-use changes on sediment load and capacity reduction of Lake Ziway, Ethiopia. Natural Resources, 11(11): 530-542. https://doi.org/10.4236/nr.2020.1111031
[8] Dharmayasa, I.G.N.P., Surakit, K., Jindal, R., Thongdara, R. (2022). Investigation on impact of changes in land cover patterns on surface runoff in Ayung Watershed, Bali, Indonesia using geographic information system. Environment and Natural Resources Journal, 20(2): 168-178. https://doi.org/10.32526/ennrj/20/202100161
[9] Jiao, Y., Zhu, G., Lu, S., Ye, L., Qiu, D., Meng, G., Wang, Q., Li, R., Chen, L., Wang, Y., Si, D., Li, W. (2024). The cooling effect of Oasis Reservoir-Riparian Forest Systems in Arid Regions. Water Resources Research, 60(10): e2024WR038301. https://doi.org/10.1029/2024WR038301
[10] Salamanca, A.M., Nugroho, A., Osbeck, M., Bharwani, S., Dwisasanti, N. (2015). Managing a living cultural landscape: Bali’s subaks and the UNESCO World Heritage Site. Stockholm Environment Institute, Project Report 2015-05.
[11] Yuniti, I.G.D.A., Purba, J.H., Sasmita, N., Komara, L. L., Olviana, T., Kartika, I.M. (2022). Balinese traditional agroforestry as base of watershed conservation. Journal of Applied Agricultural Science and Technology, 6(1): 49-60. https://doi.org/10.55043/jaast.v6i1.54
[12] Andre, N., Santosa, Y., Hikmat, A. (2022). Estimation of diversity and potential utilization of agroforestry plants in Batara Village, Timor-Leste. Jurnal Pengelolaan Sumberdaya Alam dan Lingkungan, 12(4): 609-621. https://doi.org/10.29244/jpsl.12.4.609-621
[13] Nurkholis, S., Santosa, Y., Hilwan, I. (2023). Determination of optimal sample plot dimension for measuring species diversity of plants in beach forest ecosystem. Jurnal Ilmu Kehutanan, 17(1): 56-66. https://doi.org/10.22146/jik.v17i1.5316
[14] Ma, M., Wang, Q. (2022). Assessment and forecast of green total factor energy efficiency in the yellow river basin—A perspective distinguishing the upper, middle and lower stream. Sustainability, 14(5): 2506. https://doi.org/10.3390/su14052506
[15] Montagnini, F., Nair, P.K.R. (2004). Carbon sequestration: An underexploited environmental benefit of agroforestry systems. New Vistas n Agrofor, 61: 281-295.
[16] Jose, S. (2009). Agroforestry for ecosystem services and environmental benefits: An overview. Agroforestry Systems, 76(1): 1-10. https://doi.org/10.1007/s10457-009-9229-7
[17] Sun, W., Ren, C. (2021). The impact of energy consumption structure on China’s carbon emissions: Taking the Shannon–Wiener index as a new indicator. Energy Reports, 7: 2605-2614. https://doi.org/10.1016/j.egyr.2021.04.061
[18] Gao, W., Gao, X.H., An, W. (2025). Diversity, virulence, and antimicrobial profiles of Aeromonas spp. in aquaculture products: A decade-long study in Eastern China (2012–2022). International Journal of Design & Nature and Ecodynamics, 20(5): 967-978. https://doi.org/10.18280/ijdne.200502
[19] Luo, Y., Gong, Y. (2023). α diversity of desert shrub communities and its relationship with climatic factors in Xinjiang. Forests, 14(2): 178. https://doi.org/10.3390/f14020178
[20] Sandoya, V., Saura-Mas, S., Granzow-de la Cerda, I., Arellano, G., Macía, M.J., Tello, J.S., Lloret, F. (2021). Contribution of species abundance and frequency to aboveground forest biomass along an Andean elevation gradient. Forest Ecology and Management, 479: 118549. https://doi.org/10.1016/j.foreco.2020.118549
[21] Brown, S. (1997). Estimating Biomass and Biomass Change of Tropical Forests: A Primer. Food and Agriculture Organization of the United Nations, Rome.
[22] Wickham, H. (2016). ggplot2 - Elegant Graphics for Data Analysis (2nd Edition). Journal of Statistical Software, 77(2): 1-3. https://doi.org/10.18637/jss.v077.b02
[23] The R Project for Statistical Computing. R Core Team. https://www.r-project.org/.
[24] Bunn, A.G. (2008). A dendrochronology program library in R (dplR). Dendrochronologia, 26(2): 115-124. https://doi.org/10.1016/j.dendro.2008.01.002
[25] Ickowitz, A., McMullin, S., Rosenstock, T., Dawson, I., et al. (2022). Transforming food systems with trees and forests. The Lancet Planetary Health, 6(7): e632-e639. https://doi.org/10.1016/S2542-5196(22)00091-2
[26] Kambe, I., Kholil, A.Y., Dyanasari, D. (2025). Analisis Produk Alpukat (Persea americana) di Indonesia. Doctoral dissertation, Fakultas Pertanian Universitas Tribhuwana Tunggadewi.
[27] Sujarwo, W., Caneva, G., Zuccarello, V. (2019). Patterns of plant use in religious offerings in Bali (Indonesia). Acta Botanica Brasilica, 34(1): 40-53. https://doi.org/10.1590/0102-33062019abb0110
[28] Chi, S.Y., Chang, C.C., Chien, L.H. (2025). Global tangerine trade market: Revealed competitiveness and market powers. Economies, 13(7): 203. https://doi.org/10.3390/economies13070203
[29] Devi, M., Ghatani, K. (2022). The use of coconut in rituals and food preparations in India: A review. Journal of Ethnic Foods, 9(1): 37. https://doi.org/10.1186/s42779-022-00150-7
[30] Yasin, G., Farrakh Nawaz, M., Zubair, M., Qadir, I., Saleem, A.R., Ijaz, M., Gul, S., Amjad Bashir, M., Rehim, A., Rahman, S.U., Du, Z. (2021). Assessing the contribution of citrus orchards in climate change mitigation through carbon sequestration in Sargodha District, Pakistan. Sustainability, 13(22): 12412. https://doi.org/10.3390/su132212412
[31] Islamy, R.A., Hasan, V., Kilawati, Y., Maimunah, Y., Mamat, N., Kamarudin, A.S. (2024). Water hyacinth (Pontederia crassipes) bloom in Bengawan Solo River, Indonesia: An aquatic physicochemical and biology perspective. International Journal of Conservation Science, 15(4): 1885-1898. https://doi.org/10.36868/IJCS.2024.04.19
[32] Singh, W.R., Barman, S., Tirkey, G. (2021). Morphometric analysis and watershed prioritization in relation to soil erosion in Dudhnai Watershed. Applied Water Science, 11(9): 151. https://doi.org/10.1007/s13201-021-01483-5
[33] Narendra, B.H., Siregar, C.A., Dharmawan, I.W.S., Sukmana, A., Pratiwi, Pramono, I.B., Basuki, T.M., Nugroho, H.Y.S.H., Supangat, A.B., Purwanto, Setiawan, O., Nandini, R., Ulya, N.A., Arifanti, V.B., Yuwati, T. W. (2021). A review on sustainability of watershed management in Indonesia. Sustainability, 13(19): 11125. https://doi.org/10.3390/su131911125
[34] Riley, W.J., Mekonnen, Z.A., Tang, J., Zhu, Q., Bouskill, N.J., Grant, R.F. (2021). Non-growing season plant nutrient uptake controls Arctic tundra vegetation composition under future climate. Environmental Research Letters, 16(7): 074047. https://doi.org/10.1088/1748-9326/ac0e63
[35] Junk, W.J., Piedade, M.T.F., Schöngart, J., da Cunha, C.N., Goncalves, S.R.A., Wantzen, K.M., Wittmann, F. (2024). Riparian wetlands of low-order streams in Brazil: Extent, hydrology, vegetation cover, interactions with streams and uplands, and threats. Hydrobiologia, 851(7): 1657-1678. https://doi.org/10.1007/s10750-022-05056-8
[36] Li, X., Cheng, G., Fu, B., Xia, J., et al. (2022). Linking critical zone with watershed science: The example of the Heihe River basin. Earth's Future, 10(11): e2022EF002966. https://doi.org/10.1029/2022EF002966
[37] Han, Z., Song, W. (2022). Interannual trends of vegetation and responses to climate change and human activities in the Great Mekong Subregion. Global Ecology and Conservation, 38: e02215. https://doi.org/10.1016/j.gecco.2022.e02215
[38] de Souza, S.D.N.G., Batista, D.M., Quaresma, A.C., Costa, A.L., et al. (2024). Soil flooding filters evolutionary lineages of tree communities in Amazonian riparian forests. Ecology and Evolution, 14(7): e11635. https://doi.org/10.1002/ece3.11635
[39] Natawijaya, D., Yulianto, Y., Hodiyah, I., Manik, V.T., Meylani, V. (2022). Inoculation by mycorrhizal on combinations of planting media and host plant types and their effect on plant vegetative growth. International Journal of Design & Nature and Ecodynamics, 17(6): 921-927. https://doi.org/10.18280/ijdne.170613
[40] Barbault, N., Dupraz, C., Lauri, P.E., Gosme, M. (2024). Insights into fruit tree models relevant to simulate fruit tree-based agroforestry systems. Agroforestry Systems, 98(4): 817-835. https://doi.org/10.1007/s10457-024-00953-4
[41] Šerá, B. (2021). Tree regeneration by seeds in natural forests. Forests, 12(10): 1346. https://doi.org/10.3390/f12101346
[42] Kim, M., Lee, S., Lee, S., Yi, K., Kim, H.S., Chung, S., Kim, H.S., Yoon, T.K. (2022). Seed dispersal models for natural regeneration: A review and prospects. Forests, 13(5): 659. https://doi.org/10.3390/f13050659
[43] Blancas, J., Casas, A., Pérez-Salicrup, D., Caballero, J., Vega, E. (2013). Ecological and socio-cultural factors influencing plant management in Náhuatl communities of the Tehuacán Valley, Mexico. Journal of Ethnobiology and Ethnomedicine, 9(1): 39. https://doi.org/10.1186/1746-4269-9-39