Phenotypic and Molecular Detection of Multidrug-Resistant Pseudomonas aeruginosa Isolated from Urinary Tract Infection Cases in Mosul, Iraq

Roaa A. Thamer![]() | Ahmed H. Hammo
| Ahmed H. Hammo![]() | Suhail J. Fadhil
| Suhail J. Fadhil![]() | Neam M. Alhafidh
| Neam M. Alhafidh![]() | Amera M. Al-Rawi
| Amera M. Al-Rawi![]() | Ali M. Saadi*
| Ali M. Saadi*![]() | Shihab A. Al-Bajari
| Shihab A. Al-Bajari![]()
© 2025 The authors. This article is published by IIETA and is licensed under the CC BY 4.0 license (http://creativecommons.org/licenses/by/4.0/).
OPEN ACCESS
Pseudomonas aeruginosa (P. aeruginosa) is an opportunistic pathogenic bacteria that stain gram-negative and have a significant role in infections associated with healthcare. They are responsible for a range of significant infections, like skin, urinary tract, soft tissue, respiratory tract, bone, joint, systemic, and bloodstream infections. Antibiotic resistance of P. aeruginosa represents an important challenge, resulting from adaptive, acquired, and intrinsic mechanisms. This study targeted the investigation of presence of virulence determinants in P. aeruginosa phenotypically and genotypically, 95 samples delivered from patients at Alsalam Hospital, Ibn Sina Hospital in Mosul City, five isolates were P. aeruginosa, identification of bacteria and susceptibility to antibiotics were performed using Vitek compact system, molecular diagnosis was performed using 16s rRNA sequencing, and finally the biofilm formation ability of isolates was investigated by tube method. The NCBI site reported the two most antibiotic-resistant isolates, then compared them with the standard and global isolates, and later, the evolutionary relationship was determined through a phylogenetic tree. P. aeruginosa was isolated from five samples, where each isolate was able to form biofilm and showed resistance to a variety of antibiotics. Two isolates were reported by NCBI with the names AHRA931 and AHRA932 as newly discovered isolates.
MDR, P. aeruginosa, antibiotic resistance, biofilm, PCR, Vitek, phylogenetic tree
Pseudomonas aeruginosa (P. aeruginosa) is an opportunistic pathogenic bacteria that stain gram-negative and have a significant role in infections associated with healthcare. They are responsible for a range of significant infections, like skin, urinary tract, soft tissue, respiratory tract, bone, joint, systemic, and bloodstream infections, especially in patients with weakened immunity (burns, cancer, AIDS), or cystic fibrosis (CF) [1-3]. High levels of morbidity and mortality are caused by bacteremia and pneumonia, especially in hospitalized patients. CF patients are suffering from chronic infections of the lungs that are caused by the capability of P. aeruginosa to construct mucoid biofilms, which cover the bacteria from drugs and immunity, leading to prolonged complications [4, 5].
P. aeruginosa succeeds in different environments, such as sinks and hospitals, and ventilators, in addition to natural habitats such as water and soil [6, 7]. The huge genome of these bacteria facilitates their adaptability of these bacteria, enabling survival across niches and encoding metabolic pathways [8, 9].
Antibiotic resistance of P. aeruginosa represents an important challenge, resulting from adaptive, acquired, and intrinsic mechanisms. The reduced permeability of the outer membrane and efflux pumps is responsible for the intrinsic resistance of this bacterium [10, 11]. Horizontal gene transfer is responsible for the acquired resistance, where resistance genes of plasmids, integrons, or transposons are transferred to this bacterium [12]. Adaptive resistance, usually resulting from biofilm formation, arises during treatment, as biofilms reduce the penetration of antibiotics and accommodate persistent cells. The recurrence of infections and chronicity of P. aeruginosa are contributed to by these factors [13].
The pathogenicity is controlled by a variety of virulence factors that assist colonization, escape from the immune system, and damage to tissues [14]. Secreted materials like toxins, enzymes, and pigments have crucial roles in the destruction of tissues and suppression of immunity [15]. The type III secretion system (T3SS) injects effector proteins like exoenzyme T (exoT), exoenzyme S (exoS), exoenzyme Y (exoY), and exoenzyme U (exoU) into host cells, disrupting processes of the cells and promoting bacterial invasion [16].
Pyoverdine and pyocyanin are pigments that further enhance virulence. Pyocyanin generates reactive oxygen species, resulting in oxidative damage to tissue, especially in lungs with CF. Pyoverdine, a siderophore, allows iron acquisition that is necessary for growth and infection [17]. Antimicrobial resistance and virulence together represent an enormous challenge, especially in multidrug-resistant (MDR) strains[18].
The combination of genotype with phenotype in P. aeruginosa is useful in understanding the pathogenic mechanisms of this bacterium, such as the formation of biofilm and the production of toxins. This approach explains the virulence and resistance at the molecular level, providing insight into clinical results and leading to the development of novel therapeutics [19, 20]. Anti-virulence strategies against specific toxins or biofilms give hope in relieving infections without the development of resistance [21].
This study targeted the investigation of the presence of virulence determinants in P. aeruginosa phenotypically and genotypically, and their relationship with resistance to antibiotics in both MDR and non-MDR species by linking phenotypic traits, like the formation of biofilm, with phylogenetic analysis. This work gives novel insights into the relation between virulence and resistance, thereby providing a comprehensive analysis to inform therapeutic plans.
The current study included 95 urine samples taken from incoming and inpatient patients at Al Salam Hospital and Ibn Sina Hospital, who were suffering from chronic and recurrent urinary tract diseases, including women, men, and children, aged between 9 and 70 years, over a period of five months.
Urine samples were collected using sterile plastic containers with tight lids. Approximately 10 ml of Medstrem was collected from each patient, following the patient's instructions on how to give the sample. Each sample was divided into two parts: the first was subjected to microscopic examination, and the second was placed in sterile tubes for culture on culture media, including blood agar, chocolate agar, and MacConkey agar. Subculture was performed in the media, which showed a positive cultural result.
Microscopic examination: Microscopic examination was performed using Gram stain to confirm the type of isolates. Staining was performed using Gram stain according to the manufacturer's instructions.
Biochemical test: Biochemical tests, including oxidase, catalase, and coagulase, were performed to confirm the identification of bacteria.
Vitek system: Bacteria were prepared for examination and diagnosis using the Vitek technique. A 0.5% bacterial suspension was prepared, and an identification card ID was used for this purpose. ID cards were placed in the device. After the examination period had elapsed, an accurate diagnosis was obtained for each bacterium.
Measuring bacterial resistance to antibiotics: The Vitek technique was used to diagnose bacterial Resistance to antibiotics using the AST card.
2.1 AST evaluation by the Vitek system
The Vitek Antimicrobial Susceptibility Testing (AST) Card system is based on the principle of measuring bacterial growth in the presence of different concentrations of antibiotics. It follows the guidelines of the Clinical and Laboratory Standards Institute (CLSI). The AST card is loaded with a sample of suspended bacteria and contains small wells with different concentrations of antibiotics. The system monitors bacterial growth using optical density or color change resulting from bacterial metabolic activity. The presence of each antibiotic is compared to reference readings to determine minimum inhibitory concentration (MIC) and classify bacteria as susceptible, intermediate, or resistant according to the reference standard [22]. P. aeruginosa isolates, resistant to more than three antibiotic classes, were classified as multidrug-resistant (MDR). P. aeruginosa isolates were tested against the following antibiotics: imipenem (IMI), amikacin (AK), tobramycin (TOB), ceftazidime (CAZ), ciprofloxacin (CIP), aztreonam (ATM), gentamicin (GEN), meropenem (MEM), levofloxacin (LEV), and norfloxacin (NOR).
2.2 Investigation of the ability of the bacterial strain to form biofilm
Tube Method: Pseudomonas aeruginosa was cultured in nutrient broth for 48 hours at 37℃. After incubation, the broth was carefully discarded, and the tubes were gently rinsed with buffered saline solution (BSS) to remove any non-adherent bacterial cells. The tubes were then inverted and left to air dry. Subsequently, each tube was filled with methylene blue stain and incubated for 20 minutes. The stain was then discarded, and the tubes were gently rinsed again with BSS to remove excess dye. Finally, the tubes were placed upside down and allowed to dry completely [23].
2.3 PCR amplification 16S rRNA genes
DNA was extracted using a kit provided by Geneaid. The extraction and purification steps were carried out according to the protocol.
24-hour-old cultures of Pseudomonas aeruginosa were used and grown in an N.A. medium. The cells were transferred to a 1.5 cm3 Eppendorf tube, and the bacterial cells were sedimented by centrifugation at 15,000 rpm for 5 minutes. The filtrate was discarded.
The precipitate was suspended by adding 200 µL of Gram Buffer and 200 µL of Lysozyme, mixing using a Vortex for 10 seconds, and incubating at 37℃ for 30 minutes.
20 µL of Protenase K was added, mixing using a Vortex for 10 seconds, and incubating at 60℃ for 10 minutes.
200 µL of GN Buffer was added and mixed using a Vortex. Vortex for 10 seconds and place in a water bath at 70℃ for 10 minutes. Add 200 µL of absolute ethanol after the incubation period is complete. Mix using a Vortex for 10 seconds. Place the GD Column tube inside a 2 cm3 collection tube. Transfer the contents to a GD Column tube and place it in the centrifuge. Then, replace the collection tube with another one. Add 400 µL of Washing Buffer and centrifuge at 15,000 rpm for 1 minute. Discard the liquid in the collection tube and return the tubes to the centrifuge at 15,000 rpm for 3 minutes. Add 60 µL of Elution Buffer preheated to 60℃ and incubate at room temperature for 3 minutes. At 15,000 rpm for half a minute, and store at -20℃ until ready to use [24].
The concentration and purity of DNA were measured using the BioDrop nanodrop device from the British company. The device was zeroed using TE solution, and 1 microliter of extracted DNA was added to the device to estimate the concentration in nanograms/microliter. The test was conducted at the Northern Technical University Research Center [25].
Polymerase chain reaction (PCR) was performed in a final reaction volume of 20 μL using GoTaq G2 Green Master Mix (Promega, USA). To amplify the full-length 16S rRNA gene, the universal primers 27F (5′-AGAGTTTGATCMTGGCTCAG-3′) and 1522R (5′-AAGGAGGTGATCCARCCGCA-3′) were used. Each primer was added at a final concentration of 1 μM, and 100 ng of template DNA was included in the reaction, following the manufacturer’s protocol. The PCR thermal cycling conditions were as follows: initial denaturation at 95℃ for 3 minutes, followed by 30 cycles of denaturation at 95℃ for 30 seconds, annealing at 55℃ for 30 seconds, and extension at 72℃ for 1 minute. A final extension step was performed at 72℃ for 5 minutes. The resulting PCR products were separated by electrophoresis on 2% agarose gels [26, 27], and the obtained sequences were used to identify homology between the input sequences and the reference sequences available in the NCBI GenBank database. Additionally, the phylogenetic tree was generated using the MEGA-11K software through 100X bootstrap analysis [28, 29].
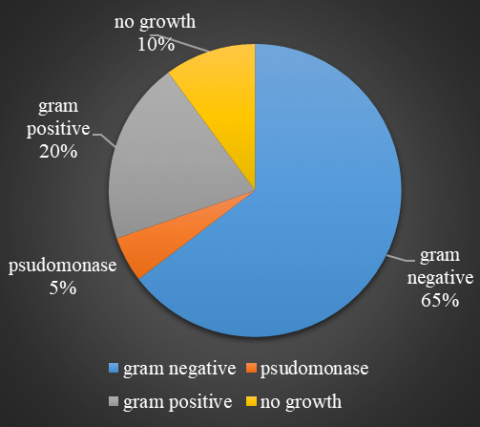
The study result showed that about 65% isolates of the total number of isolates were gram-negative bacteria, including Pseudomonas spp., while gram-positive bacteria represented 20% isolates of the total number of isolates, and 10% no growth (Figure 1).
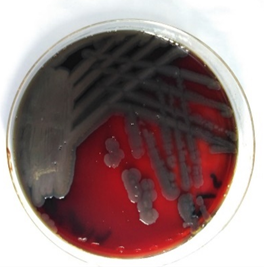
Figure 1. P.aeruginosa on BA
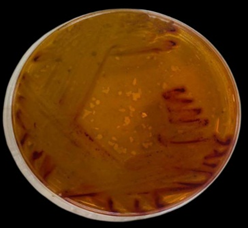
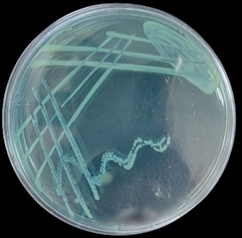
Five isolates of Pseudomonas spp. show growth on Blood agar with black colonies, then subcultured on MacConkey agar and nutrient agar, giving orange and greenish color and pale, non-lactose fermenter colonies on nutrient and MacConkey agar, respectively (Figures 2-4).
Figure 2. Culture result rate
Figure 3. P. aeruginosa on MacConky
Figure 4. P. aeruginosa on N.A
Microscopic examination showed Gram-negative bacilli (Figure 5), the manual biochemical tests exhibited oxidase-positive aerobic bacteria, and the diagnosis was confirmed by using the Vitek 2 system. The findings of our study are consistent with previous research conducted by other investigators. The high rate of bacterial growth on various agar media agreed with the results of research [30], which demonstrated that these media are effective for isolating and identifying Gram-negative bacteria from UTI infection, particularly those associated with nosocomial infections. Similarly, the observation that about 20% of the samples exhibited growth only on blood and chocolate agar agrees with the results of the study [31], which noted that certain bacterial species may not grow on MacConkey agar due to its selective properties.
Utilizing microscopic examination and manual biochemical tests to confirm the diagnosis is a well-established approach that has been supported by various studies [32]. The results of the microscopic examination showed that the bacteria were Gram-negative bacilli, as shown in Figure 5. The biochemical examinations showed that the bacteria were positive for the oxidase test, which indicates that they contain the cytochrome oxidase enzyme, and positive for the catalase test, as they produce hydrogen peroxide, and negative for the coagulase test, which confirms that they are Gram-negative bacteria.
Figure 5. Microscopic gram-negative Pseudomonas 100x
However, the application of automated identification systems such as the Vitek 2 compact system represents a more modern and efficient method for bacterial identification, offering rapid and accurate results.
Our study agreed with the study [33], which isolated various bacteria, including P. aeruginosa, from urine samples of adults.
3.1 Results of the Vitek 2 compact system
The Vitek 2 compact system was used for the Identification of P. aeruginosa. The probability was 95% - 98% for five isolates identified as P. aeruginosa. Figure 6 shows the isolates obtained from cultures and identified by the Vitek 2 system in detail.
Figure 6. Identified by Vitek 2
3.2 AST evaluation by the Vitek system
The results in Table 1 of measuring antibiotic resistance using AST card Vitek system showed that the Pseudomonas isolates were 3 of 5 resistant to CAZ, 2 of 5 resistant to MEM, 1 of 5 resistant to ATM, 2 of 5 resistant to IMI, 2 of 5 resistant to GM, 2 of 5 resistant to AK, 1 from 5 resistant to TOB, 2 from 5 resistant to CIP, 2 of 5 resistant to NOR, and 2 of 5 resistant to LEV. This result is consistent with studies [34, 35], which conducted research on Pseudomonas aeruginosa and investigated its resistance to antibiotics extensively. P. Aeruginosa isolates, resistant to more than three antibiotic classes, were classified as multidrug-resistant (MDR).
Table 1. Antibiotic resistance of isolates
|
|
LEV |
NOR |
CIP |
TOB |
AK |
GM |
IMI |
ATM |
MEM |
CAZ |
|
1 |
R |
I |
R |
R |
R |
R |
I |
R |
I |
R |
|
2 |
R |
I |
R |
S |
I |
R |
R |
S |
I |
R |
|
3 |
I |
I |
S |
S |
I |
S |
R |
I |
R |
R |
|
4 |
I |
R |
S |
I |
R |
S |
I |
I |
R |
I |
|
5 |
S |
R |
S |
I |
S |
S |
I |
I |
I |
S |
P. Aeruginosa isolates were tested against the following antibiotic disks: imipenem (IMI), amikacin (AK), aztreonam (ATM), ceftazidime (CAZ), ciprofloxacin (CIP), tobramycin (TOB), gentamicin (GEN), meropenem (MEM), levofloxacin (LEV), and norfloxacin (NOR) (Table 2).
Table 2. Antibiotic class use for detecting resistance of bacteria
|
Abbreviation |
Full Name |
Antibiotic Class |
|
CAZ |
Ceftazidime |
β-lactam Cephalosporin [3rd generation] |
|
MEM |
Merpenem |
β-lactam -Carbapenem |
|
ATM |
Azteronam |
β-lactam- Monobactam |
|
IMI |
Impenem |
β-lactam- Carbapenem |
|
GM |
Gentamicin |
Aminoglycoside |
|
AK |
Amikacin |
Aminoglycoside |
|
TOB |
Tobramycin |
Aminoglycoside |
|
CIP |
Ciprofloxacin |
Fluoroquinolone |
|
NOR |
Norfloxacin |
Fluoroquinolone |
|
LEV |
Levofloxacin |
Fluoroquinolone |
All five isolates (100%) were resistant to ceftazidime (CAZ), a third-generation cephalosporin commonly used to treat P. aeruginosa infections. This complete resistance suggests the presence of β-lactamase enzymes or active efflux mechanisms, which are well-documented contributors to β-lactam resistance. Resistance to carbapenems was also notable, with 2 out of 5 isolates (40%) resistant to both meropenem (MEM) and imipenem (IMI). Carbapenem resistance in P. aeruginosa is often linked to the production of metallo-β-lactamases (MBLs) such as VIM and IMI, or mutations leading to porin loss. These findings are consistent with research [36], which reported widespread carbapenem resistance in clinical isolates from the Middle East, emphasizing the role of mobile genetic elements in spreading resistance. Aztreonam (ATM) resistance was observed in 1 isolate (20%), indicating relatively preserved activity. However, resistance to monobactams like ATM can still emerge via β-lactamase production, particularly in MDR strains. Aminoglycoside resistance was variable: gentamicin (GEN) and tobramycin (TOB) showed resistance in 2 isolates each (40%), while amikacin (AK) had the highest resistance rate at 4 out of 5 isolates (80%). This pattern aligns with research [37], which found high levels of aminoglycoside resistance in Egyptian P. aeruginosa isolates.
Fluoroquinolone resistance was also present, with 2 isolates resistant to ciprofloxacin (CIP) and levofloxacin (LEV), and 1 isolate resistant to norfloxacin (NOR). These rates reflect the global trend of increasing fluoroquinolone resistance, enhanced by efflux pump activity. Gasink et al. [38] highlighted similar mechanisms in their study, noting the clinical impact of fluoroquinolone resistance on treatment outcomes. According to international standards, P. aeruginosa isolates resistant to three or more antibiotic classes are classified as multidrug-resistant (MDR). Several isolates in this study meet that criterion, underscoring the urgent need for antimicrobial stewardship and infection control measures.
3.3 Result of biofilm
The assessment of biofilm formation by the bacterial isolates gives positive results, shown in Figure 7. Tube B demonstrated a positive result, as evidenced by the presence of a methylene blue-stained film, indicating the capacity of the bacteria to adhere to and establish biofilms on abiotic surfaces. This finding highlights the inherent ability of these isolates to form protective biofilm structures, which are critical for their persistence and resistance in non-living environments. The results of the tube method assay for detecting the ability of Pseudomonas isolates to form biofilms revealed that all isolates were positive. This indicates that the isolates possess a strong capability to produce extracellular polymeric substances (EPS), which are the main structural components of the biofilm matrix. This property represents one of the most important virulence factors of Pseudomonas, as it enables the bacteria to adhere to both biotic and abiotic surfaces and to establish highly organized and protected cellular communities. Bacteria within the biofilm exhibit a significantly higher resistance to antibiotics compared to planktonic cells. This enhanced resistance is attributed to several mechanisms, including the physical barrier formed by the EPS matrix that limits antibiotic penetration, the reduced metabolic activity of the embedded cells, which decreases the effectiveness of drugs targeting actively dividing cells, and the presence of dormant “persister” cells that survive antimicrobial treatments. Moreover, biofilm formation provides Pseudomonas with protection against various environmental stresses and host immune defenses. The biofilm shields the bacteria from desiccation, pH fluctuations, and oxidative stress, while also impairing the efficiency of phagocytosis and other immune responses. Consequently, infections caused by Pseudomonas are often chronic and difficult to eradicate, particularly in urinary tract infections, respiratory infections, and nosocomial infections associated with medical devices such as catheters and ventilator tubes. Therefore, the positive biofilm formation observed in all Pseudomonas isolates strongly indicates their high virulence and antimicrobial resistance potential [20]. Several studies have reported a strong correlation between biofilm production and increased antibiotic resistance in P. aeruginosa isolates. For instance, Shrestha et al. [39] demonstrated that biofilm-forming isolates showed significantly higher resistance to multiple antibiotics compared to non-biofilm producers. Similarly, Tuon et al. [40] highlighted the role of biofilm in the persistence and pathogenicity of P. aeruginosa, particularly in nosocomial infections. The detection of biofilm production in this study supports these findings and suggests a potential mechanism contributing to the observed resistance patterns. The positive result for biofilm formation in Pseudomonas aeruginosa using the tube method indicates a significant health concern. Biofilm formation allows the bacteria to adhere to surfaces and produce extracellular polymeric substances (EPS), creating a protective matrix that enhances bacterial survival. This phenotype is clinically important because biofilm-producing bacteria are substantially more resistant to antibiotics and host immune responses compared to their planktonic (free-floating) counterparts. From a public health perspective, biofilm-forming P. aeruginosa is associated with chronic and persistent infections, especially in hospital settings. These include ventilator-associated pneumonia, catheter-related bloodstream infections, chronic wound infections, and complications in cystic fibrosis patients. The biofilm structure limits antibiotic penetration, facilitates horizontal gene transfer of resistance traits, and enables bacteria to enter a dormant state, making them harder to eradicate. Several studies have highlighted the danger of biofilm-associated infections. According to Harms et al. [41], biofilms contribute to up to 80% of chronic infections and are a major factor in treatment failure. Pang et al. [42] emphasized that P. aeruginosa biofilms are linked to increased morbidity and longer hospital stays due to their resilience and adaptability. The ability of P. aeruginosa to form biofilms significantly increases its pathogenic potential and complicates clinical management, underscoring the need for early detection, targeted therapies, and strict infection control measures [43].
Figure 7. The result of the biofilm formation assay, where tube B represents the positive result, showing visible biofilm formation, while tube A serves as the control
3.4 Result of PCR amplification of 16S rRNA genes
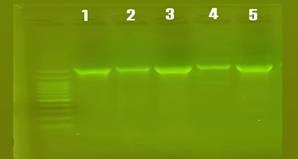
The result of amplification of the 16S rRNA gene from the genomic DNA isolated showed in Figure 8 of five P. aeruginosa isolates were identified as antibiotic-resistant Pseudomonas isolates. 16S rRNA gene analysis plays a crucial role in confirming bacterial identity and assessing the evolutionary relationships among isolates. This molecular approach enables researchers to determine whether resistant strains are genetically related—indicating possible clonal spread—or whether resistance has emerged independently in genetically distinct strains. Understanding these relationships is essential for monitoring the dissemination of resistance, identifying potential sources of infection, and guiding public health and infection control strategies.
Figure 8. PCR-amplified product 16S rRNA, the result of electrophoresis of the amplification products in a 1% agarose gel at 65 volts for an hour and a half, using the DNA ladder size indicator starting from 100 base pairs. The remaining lanes are the gene products of the isolates that showed a size of 1495 bp
These relationships are based on 16S rRNA sequences, analyzed using MEGA-11 software with a scale bar of 0.005.
The bootstrap test results (100 replicates) indicate the percentage of replicate trees where the associated strains are grouped, displayed next to the corresponding branches [44].
Our selection of these two isolates was due to their highest antibiotic resistance when an antibiotic sensitivity test was performed by using the Vitek AST card.
3.5 Phylogenetic tree
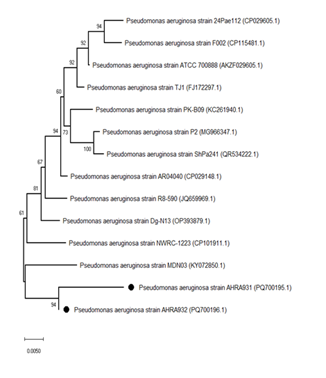
We compared the gene sequences of our isolates (1) AHRA931, (2) AHRA932, we compared with the sequences of the global isolates and the standard isolates P. aeruginosa strain ATCC 700888 [AKZF029605] in order to find the extent of similarity of these isolates in the gene sequences shown in Table 3. Through this, we can determine the extent of the bacterial epidemic. Our isolates, which were reported by the NCBI with the names AHRA931 and AHRA932, “isolates with multiple resistance to antibiotic treatment,” showed multiple resistance to antibiotics MDR. The phylogenetic tree diagram shows the extent of the relationship between the isolates (Figure 9). AHRA931 [PQ700195.1] and AHRA932 [PQ700196.1] with a very high genetic similarity of approximately 99–100% between them. A comparison with other Pseudomonas aeruginosa strains in the GenBank database yielded similarities that ranged from 96.2% to 97.85% for the following reference strains: 24Pae112 (96.2%), F002 (96.48%), ATCC 700888 (96.8%), TJ1 (97.2%), PK-B09 (97.34%), P2 (97.66%), and ShPa241 (97.85%). These results are in confirmation that both AHRA931 and AHRA932 are of the same bacterial species, Pseudomonas aeruginosa, with minor genetic variations compared to other reference isolates; this suggests that they represent different local variants within the same species (Table 3).
Figure 9. The Neighbor-Joining phylogenetic trees illustrate the relationships between Pseudomonas aeruginosa strains AHRA931 [PQ700195.1] and AHRA932 [PQ700196.1], represented by black circles, alongside other related Pseudomonas aeruginosa strains
Table 3. Most of the strains related to the numbers of accession that show homology with Pseudomonas aeruginosa strain AHRA931 [PQ700195.1] and Pseudomonas aeruginosa strain AHRA932 [PQ700196.1] retrieved from the NCBI database
|
Strain Name |
Accession No. |
Similarity (%) |
|
MDN03 |
KY072850.1 |
99.6 |
|
NWRC-1223 |
CP101911.1 |
99.37 |
|
Dg-N13 |
OP393879.1 |
99.2 |
|
R8-590 |
JQ659969.1 |
98.7 |
|
AR04040 |
CP029148.1 |
98.32 |
|
ShPa241 |
QR534222.1 |
97.85 |
|
P2 |
MG966347.1 |
97.66 |
|
PK-B09 |
KC261940.1 |
97.34 |
|
TJ1 |
FJ172297.1 |
97.2 |
|
ATCC 700888 |
AKZF029605.1 |
96.8 |
|
F002 |
CP115481.1 |
96.48 |
|
24Pae112 |
CP029605.1 |
96.2 |
The slight differences in similarity suggest potential genomic divergence, which could be related to developing mechanisms of antibiotic resistance. The selection of AHRA931 and AHRA932 was based on their remarkable resistance profiles, making them important candidates for further investigation.
These findings highlight the need for deep genomic and phenotypic characterization of these isolates for a better understanding of their resistance mechanisms. Such studies could contribute to the global understanding of Resistance to antibiotics in P. aeruginosa and aid in the development of more effective treatment strategies, particularly in regions where resistant strains are emerging.
High levels of antibiotic resistance shown by AHRA931 and AHRA932 were the primary reason for their selection. The phylogenetic analysis based on 16S rRNA gene sequencing provided valuable insights into the genetic relationships among the antibiotic-resistant Pseudomonas isolates. Despite phenotypic similarities in antibiotic resistance profiles, the sequence-based analysis revealed varying degrees of genetic diversity among the isolates. This indicates that resistance may not be solely due to clonal expansion of a single strain, but could also result from independent acquisition of resistance genes by genetically distinct strains [45, 46].
The clustering of some isolates within the same phylogenetic branch suggests a close evolutionary relationship, possibly pointing to a common origin or transmission route. In contrast, other isolates positioned in separate branches reflect genetic heterogeneity, emphasizing the adaptive capability of Pseudomonas species in different environments and under selective pressure from antibiotics [47]. These findings highlight the importance of molecular tools like 16S rRNA sequencing in complementing phenotypic methods, especially for epidemiological surveillance and tracking the emergence and spread of resistance.
The current study was conducted on 95 clinical urine samples obtained from patients with symptoms associated with urinary tract infection. The results showed that 85 samples (89.5%) were positive for bacterial growth, indicating a high infection rate in the sample studied. This percentage is a strong indicator of a real health problem with regard to urinary tract infections, especially when compared to previous studies that reported lower positive rates. For example, Said et al. [48] indicated that the positivity rate in urine cultures in a teaching hospital was about 65%, while another study by Abbas et al. [49] in Egypt showed a rate of about 70%, reflecting the diversity of infection rates depending on the clinical environment, the nature of the patients, and the isolation methods used.Among the isolates obtained, five were identified as belonging to the genus Pseudomonas using a series of morphological and biochemical tests. The isolates were cultured on selective and diagnostic media such as MacConkey agar, nutrient agar, and blood agar, where the isolates showed characteristics consistent with Pseudomonas aeruginosa, such as growth on MacConkey without lactose fermentation, production of green pigment on nutrient media, and Gram-negative rod shape, which supports the preliminary identification of the isolates. All isolates also gave positive results for the oxidase test, which reinforces their diagnosis as P. aeruginosa. The diagnosis was confirmed using the VITEK 2 device, which showed complete agreement with the results of conventional tests. The use of this device is an important step for diagnostic accuracy, as it relies on an extensive database of biochemical tests and metabolic patterns. Results with regard to antibiotic susceptibility testing, the five isolates showed notable resistance to several classes of antibiotics, including third-generation cephalosporins, carbapenems, and fluoroquinolones. Resistance to more than three classes of antibiotics indicates that the isolates are classified as multidrug-resistant (MDR) bacteria, which increases the risk of clinical management. These findings are consistent with those reported by Rahimzadeh et al. [50] in Iran, which documented high resistance rates in P. aeruginosa isolates from urinary tract patients, with carbapenem resistance exceeding 60%.In the same context, Nader et al. [51] from Iraq indicated that most clinical Pseudomonas isolates showed high resistance to aminoglycosides and carbapenems, in addition to a close association between resistance and biofilm formation. This is what we observed in our study, as the five isolates showed a clear tendency to form biofilms when tested using the tube method, forming a dense layer of biofilm on the tube walls, with varying degrees of intensity from moderate to strong.Biofilm formation is an important virulence trait of P. aeruginosa, as it gives it the ability to adhere to surfaces, resist antibiotics, and evade the immune system. El-Sayed and Fahmy [52] indicated that more than 80% of P. aeruginosa isolates from patients with urinary tract infections showed strong biofilm formation, and all were resistant to more than one class of antibiotics, confirming the relationship between these two characteristics, as in our results. To support the results of conventional diagnosis, molecular analysis was used by amplifying and sequencing the 16S rRNA gene, which is considered the gold standard in bacterial diagnosis. The sequencing results showed a high degree of similarity with global isolates documented in the NCBI database, and phylogenetic tree analysis allowed the construction of phylogenetic relationships between local and international isolates. This analysis showed that the study isolates belong to the same evolutionary branches that include P. aeruginosa strains isolated from hospitals in India, Turkey, and Europe, indicating the spread of a common genetic pattern.These results are consistent with those reported in the study of Al-Shimmary et al. [53], which used 16S rRNA sequencing to construct a phylogenetic tree of resistant Pseudomonas isolates. and found that there was significant genetic similarity between isolates from different countries, supporting the idea that some resistant strains have a common origin or have acquired similar characteristics due to selective pressure from antibiotics in similar clinical environments.Therefore, the results of this study highlight the importance of monitoring urinary Pseudomonas aeruginosa isolates, especially those that show multidrug resistance and biofilm formation, due to the therapeutic challenges they pose. At the same time, they emphasize the importance of integrating traditional and molecular tests to confirm the identity and identify the evolutionary relationships between isolates. One limitation of this study is that the comparatively small sample size constrains the generalizability of the results. However, the results do provide valuable preliminary evidence of attributes of the locally prevailing MDR P. aeruginosa strains and affirm the need for future large-scale investigations.
The systematic linking of phenotypic and biochemical bacterial diagnostic results with antibiotic resistance profiling represents a crucial step in understanding the clinical and epidemiological characteristics of bacterial isolates, particularly in the context of urinary tract infections caused by Pseudomonas aeruginosa. This integration not only provides a comprehensive picture of the isolated bacteria, but also helps to explain their biological behavior within the host and the effectiveness of available treatment options.Resistant bacterial isolates typically exhibit advanced virulence characteristics, most notably the ability to form biofilms, which represent one of the most prominent strategies of bacteria to resist external factors, whether antibiotics or immune responses. Morphological investigation of biofilms using tube tests or microplates contributes to a direct link between the phenotypic characteristics of bacteria and their resistance pattern. The results of this study indicate a close correlation between multidrug-resistant isolates and their ability to form biofilms, supporting previous literature that biofilm production is associated with increased resistance mechanisms such as reduced cell wall permeability, increased efflux pump activity, and the presence of persistent cells. These findings reinforce the hypothesis that the integration of these traits—drug resistance and membrane formation—is not random behavior, but rather an adaptive evolution driven by selection pressure in the clinical environment. Hence, the importance of adding 16S rRNA molecular analysis to this study, which provided an additional level of understanding by comparing locally isolated strains with global standard isolates. Phylogenetic analysis showed that locally isolated strains share close evolutionary relationships with globally known isolates from hospitals in Asia, Europe, and North America, indicating the presence of widespread strains with evolutionarily conserved characteristics in terms of resistance and virulence. This genetic similarity between local and global isolates supports the hypothesis of resistance strain transmission through multiple channels, such as medical travel, spread within healthcare facilities, or through the exchange of resistance genes via plasmids and transferable sequences. It also highlights the epidemiological importance of monitoring local isolates within a global context, especially when these isolates exhibit complex resistance patterns and active membrane formation. Therefore, linking the results of transplantation and traditional laboratory diagnosis with the results of antibiotic resistance, through the evaluation of phenotypic behavior (such as biofilm production), and ending with molecular evolutionary analysis, represents a highly valuable integrated approach to the evaluation of bacterial isolates from a clinical and epidemiological perspective. This integration not only contributes to improved diagnostic accuracy but also paves the way for the design of evidence-based treatment protocols and surveillance strategies that seek to limit the spread of multidrug-resistant and virulent P. aeruginosa strains within healthcare facilities.
The authors would like to express their sincere gratitude to Northern Technical University, the Technical Institute of Health, and the Department of Medical Laboratory Techniques for their support.
This study was approved by the Northern Technical University as a research study using anonymous medical data, thus exempting researchers from obtaining informed consent from patients as usual.
[1] De Bentzmann, S., Plésiat, P. (2011). The Pseudomonas aeruginosa opportunistic pathogen and human infections. Environmental Microbiology, 13(7): 1655-1665. https://doi.org/10.1111/j.1462-2920.2011.02469.x
[2] Jaafar, F., Bashar, N., Alhusseini, L., Musafer, H. (2023). Infections with Pseudomonas aeruginosa in Burn Patients: The host immune response. South Asian Research Journal of Biology and Applied Biosciences, 5(2): 15-21. https://doi.org/10.1111/j.1462-2920.2011.02469.x
[3] Neu, H.C. (2020). Clinical role of Pseudomonas aeruginosa. In Glucose Nonfermenting Gram-negative Bacteria in Clinical Microbiology, pp. 83-104.
[4] Osmon, S., Ward, S., Fraser, V.J., Kollef, M.H. (2004). Hospital mortality for patients with bacteremia due to Staphylococcus aureus or Pseudomonas aeruginosa. Chest, 125(2): 607-616. https://doi.org/10.1378/chest.125.2.607
[5] Bhagirath, A.Y., Li, Y., Somayajula, D., Dadashi, M., Badr, S., Duan, K. (2016). Cystic fibrosis lung environment and Pseudomonas aeruginosa infection. BMC Pulmonary Medicine, 16(1): 174. https://doi.org/10.1186/s12890-016-0339-5
[6] Crone, S., Vives‐Flórez, M., Kvich, L., Saunders, A.M., et al. (2020). The environmental occurrence of Pseudomonas aeruginosa. Apmis, 128(3): 220-231. https://doi.org/10.1111/apm.13010
[7] Streeter, K., Katouli, M. (2016). Pseudomonas aeruginosa: A review of their pathogenesis and prevalence in clinical settings and the environment. Infection, Epidemiology and Medicine, 2(1): 25-32.
[8] Moradali, M.F., Ghods, S., Rehm, B.H. (2017). Pseudomonas aeruginosa lifestyle: A paradigm for adaptation, survival, and persistence. Frontiers in Cellular and Infection Microbiology, 7: 39. https://doi.org/10.3389/fcimb.2017.00039
[9] De Sousa, T., Hébraud, M., Dapkevicius, M.L.E., Maltez, L., Pereira, J.E., Capita, R., Alonso-Calleja, C., Igrejas, G., Poeta, P. (2021). Genomic and metabolic characteristics of the pathogenicity in Pseudomonas aeruginosa. International Journal of Molecular Sciences, 22(23): 12892. https://doi.org/10.3390/ijms222312892
[10] Yang, J., Xu, J.F., Liang, S. (2025). Antibiotic resistance in Pseudomonas aeruginosa: Mechanisms and emerging treatment. Critical Reviews in Microbiology, 51(5): 841-859. https://doi.org/10.1080/1040841X.2024.2429599
[11] Lorusso, A.B., Carrara, J.A., Barroso, C.D.N., Tuon, F.F., Faoro, H. (2022). Role of efflux pumps on antimicrobial resistance in Pseudomonas aeruginosa. International Journal of Molecular Sciences, 23(24): 15779. https://doi.org/10.3390/ijms232415779
[12] Lerminiaux, N.A., Cameron, A.D. (2019). Horizontal transfer of antibiotic resistance genes in clinical environments. Canadian Journal of Microbiology, 65(1): 34-44. https://doi.org/10.1139/cjm-2018-0275
[13] Soares, A., Alexandre, K., Etienne, M. (2020). Tolerance and persistence of Pseudomonas aeruginosa in biofilms exposed to antibiotics: Molecular mechanisms, antibiotic strategies and therapeutic perspectives. Frontiers in Microbiology, 11: 2057. https://doi.org/10.3389/fmicb.2020.02057
[14] Abdulateef, S.A., Owaif, H.A.A., Hussein, M.H. (2023). Importance of virulence factors in bacterial pathogenicity: A review. International Journal of Medical Science and Clinical Research Studies, 3(4): 765-769. https://doi.org/10.47191/ijmscrs/v3-i4-35
[15] Tam, K., Torres, V.J. (2019). Staphylococcus aureus secreted toxins and extracellular enzymes. Microbiology Spectrum, 7(2). https://doi.org/10.1128/microbiolspec.gpp3-0039-2018
[16] Hardy, K.S., Tessmer, M.H., Frank, D.W., Audia, J.P. (2021). Perspectives on the Pseudomonas aeruginosa type III secretion system effector ExoU and its subversion of the host innate immune response to infection. Toxins, 13(12): 880. https://doi.org/10.3390/toxins13120880
[17] Sahoo, S.R., Pradhan, A.K., Das, R.P., Panigrahi, L.L., Arakha, M. (2023). Pyocyanin is the microbial blue-green pigment: A review on its history, virulence, and therapeutic use. Current Bioactive Compounds, 19(6): 62-76. https://doi.org/10.2174/1573407219666221209092023
[18] Geisinger, E., Isberg, R.R. (2017). Interplay between antibiotic resistance and virulence during disease promoted by multidrug-resistant bacteria. The Journal of Infectious Diseases, 215(suppl_1): S9-S17. https://doi.org/10.1093/infdis/jiw402
[19] Rodrigues, R.L., Lima, J.L.D.C., Sena, K.X.D.F.R.D., Maciel, M.A.V. (2020). Phenotypic and genotypic analysis of biofilm production by Pseudomonas aeruginosa isolates from infection and colonization samples. Revista da Sociedade Brasileira de Medicina Tropical, 53: e20200399. https://doi.org/10.1590/0037-8682-0399-2020
[20] Fadhil, S.J., Alomari, S.M., Al-Najim, A.N., Qasinr, R.Y. (2025). Occurrence of biofilm forming and antibiotic resistance of clinical Stenotrophomonas maltophilia in Ninawa hospitals, Iraq. Microbes and Infectious Diseases, 6(2): 811-818.
[21] Fleitas Martínez, O., Cardoso, M.H., Ribeiro, S.M., Franco, O.L. (2019). Recent advances in anti-virulence therapeutic strategies with a focus on dismantling bacterial membrane microdomains, toxin neutralization, quorum-sensing interference and biofilm inhibition. Frontiers in Cellular and Infection Microbiology, 9: 74. https://doi.org/10.3389/fcimb.2019.00074
[22] Carvalhaes, C.G., Shortridge, D., Rhomberg, P., Castanheira, M. (2024). β-lactam resistant phenotypes reported by VITEK® 2 advanced expert systemTM (AES) compared to whole genome sequencing in Enterobacterales from North and Latin America. Diagnostic Microbiology and Infectious Disease, 110(1): 116358. https://doi.org/10.1016/j.diagmicrobio.2024.116358
[23] Hammo, A.H., Al-Rawi, A.M. (2021). Evaluation of the resistance of CuO-coated contact lenses to bacterial contamination. Journal of Physics: Conference Series, 1999(1): 012035. https://doi.org/10.1088/1742-6596/1999/1/012035
[24] Rahmadara, G., Hanifah, N.F., Rismayanti, R., Purwoko, D., Rochandi, A., Tajuddin, T. (2022). Comparison of DNA isolation methods that derived from leaves of a potential anti-cancer rodent tuber (Typhonium flagelliforme) plant. International Journal of Agriculture System, 10(2): 93-103. https://doi.org/10.20956/ijas.v10i2.3966
[25] Versmessen, N., Van Simaey, L., Negash, A.A., Vandekerckhove, M., Hulpiau, P., Vaneechoutte, M., Cools, P. (2024). Comparison of DeNovix, NanoDrop and Qubit for DNA quantification and impurity detection of bacterial DNA extracts. Plos One, 19(6): e0305650. https://doi.org/10.1371/journal.pone.0305650
[26] Iralu, N., Wani, S., Meghanath, D., Hamid, A. (2025). Polymerase Chain Reaction (PCR) for virus detection. In Detection of Plant Viruses: Advanced Techniques, pp. 111-113. https://doi.org/10.1007/978-1-0716-4390-7_20
[27] Chen, R., Lu, X., Li, M., Chen, G., Deng, Y., Du, F., Dong, J., Huang, X., Cui, X., Tang, Z. (2019). Polymerase chain reaction using “V” shape thermal cycling program. Theranostics, 9(6): 1572-1579. https://doi.org/10.7150/thno.31986
[28] O'Leary, N.A., Wright, M.W., Brister, J.R., Ciufo, S., et al. (2016). Reference sequence (RefSeq) database at NCBI: Current status, taxonomic expansion, and functional annotation. Nucleic Acids Research, 44(D1): D733-D745. https://doi.org/10.1093/nar/gkv1189
[29] Russo, C.A.D.M., Selvatti, A.P. (2018). Bootstrap and rogue identification tests for phylogenetic analyses. Molecular Biology and Evolution, 35(9): 2327-2333. https://doi.org/10.1093/molbev/msy118
[30] Okafor, J.U., Nwodo, U.U. (2023). Detection of multidrug-resistant RND efflux pumps and regulatory proteins in antibiotic-resistant P. aeruginosa recovered from hospital wastewater effluent in the Eastern Cape Province of South Africa. Applied Sciences, 13(20): 11241. https://doi.org/10.3390/app132011241
[31] Shaapera, A., Joshua, I.S. (2025). Antibiotics profiling of bacteria isolated from urine samples of individuals in Wukari, Taraba State. https://doi.org/10.2139/ssrn.5161256
[32] Maeda, M., Sunaga, T., Sato, M.T., Hasegawa, T., Noma, H., Ota, E. (2024). Efficacy of carbapenems and alternative antimicrobials for treating complicated urinary tract infections caused by third-generation cephalosporin-resistant gram-negative bacteria: A systematic review and meta-analysis of randomised controlled trials. Journal of Infection and Chemotherapy, 30(11): 1147-1155. https://doi.org/10.1016/j.jiac.2024.05.001
[33] Froom, P., Shimoni, Z. (2024). Laboratory tests, bacterial resistance, and treatment options in adult patients hospitalized with a suspected urinary tract infection. Diagnostics, 14(11): 1078. https://doi.org/10.3390/diagnostics14111078
[34] Habib, M.B., Shah, N.A., Amir, A., Alghamdi, H.A., Tariq, M.H., Nisa, K., Ammoun, M. (2025). Decoding MexB efflux pump genes: Structural, molecular, and phylogenetic analysis of multidrug-resistant and extensively drug-resistant Pseudomonas aeruginosa. Frontiers in Cellular and Infection Microbiology, 14: 1519737. https://doi.org/10.3389/fcimb.2024.1519737
[35] Horcajada, J.P., Montero, M., Oliver, A., Sorlí, L., Luque, S., Gómez-Zorrilla, S., Benito, N., Grau, S. (2019). Epidemiology and treatment of multidrug-resistant and extensively drug-resistant Pseudomonas aeruginosa infections. Clinical Microbiology Reviews, 32(4). https://doi.org/10.1128/cmr.00031-19
[36] Al-Orphaly, M., Hadi, H.A., Eltayeb, F.K., Al-Hail, H., Samuel, B.G., Sultan, A.A., Skariah, S. (2021). Epidemiology of multidrug-resistant Pseudomonas aeruginosa in the Middle East and North Africa Region. Msphere, 6(3). https://doi.org/10.1128/msphere.00202-21
[37] Esparragón, F.R., Martín, M.G., Lama, Z.G., Sabatelli, F.J., Junco, M.T.T. (2000). Aminoglycoside resistance mechanisms in clinical isolates of Pseudomonas aeruginosa from the Canary Islands. Zentralblatt für Bakteriologie, 289(8): 817-826. https://doi.org/10.1016/S0934-8840(00)80008-0
[38] Gasink, L.B., Fishman, N.O., Weiner, M.G., Nachamkin, I., Bilker, W.B., Lautenbach, E. (2006). Fluoroquinolone-resistant Pseudomonas aeruginosa: assessment of risk factors and clinical impact. The American Journal of Medicine, 119(6): 526-e19-526-e25. https://doi.org/10.1016/j.amjmed.2005.11.029
[39] Shrestha, R., Nayak, N., Bhatta, D.R., Hamal, D., Subramanya, S.H., Gokhale, S. (2019). Drug resistance and biofilm production among Pseudomonas aeruginosa clinical isolates in a tertiary care hospital of Nepal. Nepal Medical College Journal, 21(2): 110-116. https://doi.org/10.3126/nmcj.v21i2.25109
[40] Tuon, F.F., Dantas, L.R., Suss, P.H., Tasca Ribeiro, V.S. (2022). Pathogenesis of the Pseudomonas aeruginosa biofilm: A review. Pathogens, 11(3): 300. https://doi.org/10.3390/pathogens11030300
[41] Harms, A., Maisonneuve, E., Gerdes, K. (2016). Mechanisms of bacterial persistence during stress and antibiotic exposure. Science, 354(6318): aaf4268. https://doi.org/10.1126/science.aaf4268
[42] Pang, Z., Raudonis, R., Glick, B.R., Lin, T.J., Cheng, Z. (2019). Antibiotic resistance in Pseudomonas aeruginosa: Mechanisms and alternative therapeutic strategies. Biotechnology Advances, 37(1): 177-192. https://doi.org/10.1016/j.biotechadv.2018.11.013
[43] Sanya, D.R.A., Onésime, D., Vizzarro, G., Jacquier, N. (2023). Recent advances in therapeutic targets identification and development of treatment strategies towards Pseudomonas aeruginosa infections. BMC Microbiology, 23(1): 86. https://doi.org/10.1186/s12866-023-02832-x
[44] Ozer, B., Duran, N., Onlen, Y., Savas, L. (2012). Efflux pump genes and antimicrobial resistance of Pseudomonas aeruginosa strains isolated from lower respiratory tract infections acquired in an intensive care unit. The Journal of Antibiotics, 65(1): 9-13. https://doi.org/10.1038/ja.2011.102
[45] Nonaka, L., Inubushi, A., Shinomiya, H., Murase, M., Suzuki, S. (2010). Differences of genetic diversity and antibiotics susceptibility of Pseudomonas aeruginosa isolated from hospital, river and coastal seawater. Environmental Microbiology Reports, 2(3): 465-472. https://doi.org/10.1111/j.1758-2229.2010.00178.x
[46] Kiyaga, S., Kyany'a, C., Muraya, A.W., Smith, H.J., Mills, E.G., Kibet, C., Mboowa, G., Musila, L. (2022). Genetic diversity, distribution, and genomic characterization of antibiotic resistance and virulence of clinical Pseudomonas aeruginosa strains in Kenya. Frontiers in Microbiology, 13: 835403. https://doi.org/10.3389/fmicb.2022.835403
[47] Vaez, H., Faghri, J., Esfahani, B.N., Moghim, S., Fazeli, H., Sedighi, M., Safaei, H.G. (2015). Antibiotic resistance patterns and genetic diversity in clinical isolates of Pseudomonas aeruginosa isolated from patients of a referral hospital, Isfahan, Iran. Jundishapur Journal of Microbiology, 8(8): e20130. https://doi.org/10.5812/jjm.20130v2
[48] Said, A., El-Gamal, M.S., Abu-Elghait, M., Salem, S.S. (2021). Isolation, identification and antibiotic susceptibility pattern of urinary tract infection bacterial isolates. Lett Appl NanoBioSci, 10(4): 2820-2830. https://doi.org/10.33263/LIANBS104.28202830
[49] Abbas, H.A., El-Ganiny, A.M., Kamel, H.A. (2018). Phenotypic and genotypic detection of antibiotic resistance of Pseudomonas aeruginosa isolated from urinary tract infections. African Health Sciences, 18(1): 11-21. https://doi.org/10.4314/ahs.v18i1.3
[50] Rahimzadeh, M., Shahbazi, S., Sabzi, S., Habibi, M., Asadi Karam, M.R. (2023). Antibiotic resistance and genetic diversity among Pseudomonas aeruginosa isolated from urinary tract infections in Iran. Future Microbiology, 18(16): 1171-1183. https://doi.org/10.2217/fmb-2023-0118
[51] Nader, M.I., Kareem, A.A., Rasheed, M.N., Issa, M.A. (2017). Biofilm formation and detection of pslÁ gene in multidrug resistant Pseudomonas aeruginosa isolated from Thi-Qar, Iraq. Iraqi Journal of Biotechnology, 16(4).
[52] El-Sayed, H., Fahmy, Y. (2021). Correlation between biofilm formation and multidrug resistance in clinical isolates of Pseudomonas aeruginosa. Microbes and Infectious Diseases, 2(3): 541-549.
[53] Al-Shimmary, S.M., Mohamed, N.S., Al-Qaysi, S.A., Almohaidi, A.M.S. (2021). Phylogeny analysis of gyrB gene and 16S rRNA genes of Pseudomonas aeruginosa isolated from Iraqi Patients. Research Journal of Pharmacy and Technology, 14(5): 2517-2521.