Bioactive Coating from Melastoma malabathricum Enhances Fusarium Wilt Resistance and Plant Growth in Shallots on Peatland Soils

Dian Novriadhy*![]() | Sri Maryani
| Sri Maryani![]()
© 2025 The authors. This article is published by IIETA and is licensed under the CC BY 4.0 license (http://creativecommons.org/licenses/by/4.0/).
OPEN ACCESS
Fusarium wilt constrains shallot cultivation on peatlands, where acidic conditions favor the pathogen. This study evaluated bioactive coatings from Melastoma malabathricum (MM) and Azadirachta indica (AI) as sustainable fungicide alternatives. A completely randomized design was used with ten replicates per treatment: negative control, chlorothalonil (positive control), MM, and AI coatings. Under natural pathogen pressure, the MM coating—a unique acidic gel—significantly delayed Fusarium wilt onset until 24 days after planting (DAP) and showed the lowest disease incidence (20% vs. 40% in control). The AI coating (a neutral slurry) also showed activity (30% incidence). Furthermore, the MM treatment promoted synergistic plant growth, achieving the highest growth coefficient (0.795 from the quadratic model YBDS = 0.795(DAP)-0.015(DAP)2) and a strong positive interaction between treatment and leaf area development (β = 0.023, p < 0.001 in Generalized Linear Model analysis). A key observation was the recruitment of beneficial Trichoderma sp. by MM, suggesting a novel self-reinforcing mechanism, though this requires quantitative confirmation. Conversely, AI exhibited allelopathic effects that impeded early growth. The MM-based coating is a multifaceted technology for concurrent disease management and growth promotion in peatlands.
bioactive coating, plant growth synergy, Trichoderma recruitment, phytochemical antifungal
Shallots (Allium cepa var. aggregatum) are a strategic commodity in Indonesia, but their economic viability fluctuates due to climate change [1] and Fusarium sp. wilt disease [2]. Indonesia's shallot production in 2023 reached 1,985,233.3 tons with a very small increase of 0.14% compared to the previous year, while in the same period, the harvested area decreased by 3,301 ha [3, 4]. Yield losses due to Fusarium sp. wilt in 2024 are estimated to reach 10,012.67 tons, with the area of disease attack reaching 6,356.37 ha [5]. Not only does it damage shallot plants in the fields, but Fusarium sp. also causes 50% of harvested bulbs in the warehouse to experience root rot [6]. For these reasons, Fusarium sp. wilt control is a priority.
Previous research results show that the strategy for controlling Fusarium sp. wilt in shallots is towards an integrated approach [7]. Use of biological agents (Trichoderma) [8], selective fungicides [9], improved cultivation techniques [10], and storage innovations [11] shows varying degrees of success in controlling attacks. Recent research focuses on identifying antioxidant enzymes [12] and genetic markers for resistance to Fusarium infection [13, 14] as a basis for sustainable solutions. However, challenges such as pathogen adaptation, field efficacy, and environmental concerns of chemical inputs still require the development of novel, multi-functional solutions.
Conversely, the tolerance of shallots to abiotic stresses such as drought and saline soil under field conditions has been previously evaluated [15]. However, research on their adaptability to acidic soils—particularly peatlands—remains limited, to the best of the author's knowledge. This scarcity of studies is unsurprising, given that peatlands are primarily designated as carbon sinks and critical climate regulators, warranting restricted agricultural utilization [16]. Nevertheless, in accordance with Indonesian Government Regulation No. 57/2016, shallow peatlands (with a peat thickness of <100 cm) may be used for agricultural purposes, provided that the groundwater level is maintained at ≤ 40 cm from the surface [17]. This regulatory framework, coupled with the need for sustainable intensification, creates a niche for innovative agricultural technologies that can enhance crop resilience in these marginal soils while minimizing environmental impact. One promising avenue is the development of bioactive coatings that can deliver plant-based phytochemicals to protect bulbs and stimulate growth directly from the point of planting.
Despite their promise as a sustainable alternative to chemical treatments, current research on bioactive coatings faces significant limitations. These include a frequent narrow spectrum of action against pathogens and inconsistent performance when moving from controlled laboratory conditions to variable field environments. This area of research is also constrained by a limited understanding of precise ecological interaction and the long-term effects on soil microbiomes, alongside economic and regulatory barriers that complicate scaling and commercialization [18].
These challenges are particularly pronounced in specialized agricultural contexts. A critical literature gap exists at the intersection of botanical seed coatings and Allium crop cultivation on peatlands. To our knowledge, no prior study has developed an environment-specific bioactive coating for this purpose. This lack of research is significant, as peat soils present a unique set of abiotic and biotic stresses that generic coatings may not address. Our work is positioned to fill this specific void by leveraging local plant adaptations to create a targeted solution.
To address these limitations, this study investigates an innovative bioactive coating designed to enable sustainable shallot cultivation on peatlands. We developed a coating formulated with phytochemical extracts from peatland-abundant species, including Melastoma malabathricum (MM) –chosen for its native adaptation to acidic soils–while Azadirachta indica (AI) was included as a comparative control with well-established antifungal properties. The core innovation lies in the coating matrix, engineered to stabilize the phytochemicals and facilitate a slow release. This design is intended to provide simultaneous protection against disease and promotion of plant growth–a dual functionality that is often lacking in conventional treatments. This study sought to answer the following questions: (1) Can bioactive coating formulated from local peatland plant phytochemicals provide effective protection against Fusarium wilt and enhance growth parameters in shallots cultivated in peat soil? and (2) What are the potential mechanisms–including direct antifungal effects, modulation of the plant’s physiological responses, and interactions with the peat soil microbiome–that underpin the efficacy of these phytochemical coatings?
The study evaluated the efficacy of MM and AI leaf extracts in enhancing the resistance of A. cepa var. aggregatum against Fusarium sp. The experiment, conducted from June to August 2025 at the Sriwijaya Botanical Garden, employed a Completely Randomized Design with four treatments: 1) no treatment (negative control), 2) use of 75% chlorothalonil (positive control), 3) MM leaf extract, and 4) AI leaf extract. Each treatment was replicated ten times (N = 40 bulbs total), a sample size consistent with common replication standards in similar phytopathological screening trials. During the research period, the average atmospheric conditions had a relative humidity (RH) of 83% (74% - 93%) and temperature (T) of 28.4℃ (22.4℃ - 35.3℃) [19].
2.1 Phytochemical profiling
The phytochemical compounds profile with potential as biostimulants and antifungals was identified based on published literature and databases, referring to target species or the same genus if species-specific data were unavailable. The online reference sources/websites used for data collection were: 1) Dr. Duke’s Phytochemical and Ethnobotanical Databases [20], 2) PubChem [21], and 3) sciencedirect.com. The phytochemical compounds included in the study met the following criteria: 1) extracted from leaves, 2) water used as the solvent, and 3) sourced from the target species, or, if unavailable, phytochemicals from the same genus were utilized.
2.2 Pathogen source and inoculum conditions
No artificial inoculation was performed; instead, the experiment relied on natural pathogen pressure to simulate real-world disease dynamics. The pathogen population reflects the native field conditions under which the bioactive coatings are intended to be applied. This approach was chosen to evaluate the efficacy of the coatings under practical, farmer-relevant conditions, where multiple Fusarium strains and other soilborne microbes may coexist.
2.3 Material preparation
Preparation of experimental materials consists of five stages:
2.4 Bulb selection and treatment
A. cepa var. aggregatum bulbs of Bima Brebes type were obtained from onion seed breeders in Demak Regency, Central Java, Indonesia, in bulk weighing 1 kg with the specification of having been dried using sunlight for 14 days and having a shelf life of 8 weeks (April-June 2025). The readiness of the bulbs for planting was determined by cutting the bulbs crosswise and ensuring the presence of buds in the center of the bulb.
Bright reddish single bulbs are manually selected and then visually inspected to ensure they are free from physical damage, root rot, soft-watery bulbs, and/or hollow bulbs. A total of 40 bulbs, selected for uniformity with an overall average diameter of 12.75 ± 0.18 mm and an overall average weight of 1.76 ± 0.04 g, were used as seeds. These bulbs were then randomly divided into four treatment groups (n = 10 per group), with the initial characteristics of each group detailed in Table 1. The outermost layer of skin was peeled off cleanly (Figure 1) before receiving the treatment, as in Table 1.
Table 1. Bulb’s characteristics and treatments
|
Code |
Treatment |
Repetition |
Physical Characteristics of Seeds |
|
C |
Seeds did not receive antifungal treatment |
10 |
Diameter 13.02 ± 0.44 mm, weight 1.84 ± 0.06 g, bright purplish red color, no lesions on the bulbs, and good condition |
|
MM |
The seeds were soaked in EMM for 10 minutes, after which the entire surface was coated with MMP and MM powder. EMMST applied at 14 and 28 DAP |
10 |
Diameter 12.82 ± 0.38 mm, weight 1.88 ± 0.11 g, bright purplish red color, no lesions on the bulbs, and good condition |
|
AI |
The seeds were soaked in EAI for 10 minutes, after which the entire surface was coated with AIP and AI powder. EAIST applied at 14 and 28 DAP |
10 |
Diameter 12.48 ± 0.34 mm, weight 1.59 ± 0.07 g, bright purplish red color, no lesions on the bulbs, and good condition |
|
PL |
Seeds coated with fungicide (Chlorothalonil 75%) |
10 |
Diameter 12.67 ± 0.32 mm, weight 1.72 ± 0.04 g, bright purplish red color, no lesions on the bulbs, and good condition |
Figure 1. Initial bulb and encapsulated bulb appearance
2.5 Planting media preparation, growth conditions, and research variables
The soil uses sapric peat, which is dried in sunlight for 5 days to enhance the mobilization of metal elements and adjust the pH [24]. The sapric peat extracted at -3.1569 South Latitude; 104.5493 East Longitude in the Sriwijaya Botanical Gardens area, Indonesia. Dried sapric peat has the characteristics of 36.20% organic carbon; Nitrogen 0.78%; pH 6, and Cation Exchange Capacity (CEC) 64.58 cmol+/kg. The dried sapric peat is then sieved to improve drainage and aeration, which can increase the availability of phosphorus [25]. Dry sifted sapric peat as much as 4.2 kg/bag enriched with Nitrogen:Potassium:Kalium (NPK) = 16:16:16 fertilizer as much as 3 g/bag [26] and dolomite 25 g/bag [27], homogenized, became planting media (PM).
The bulb is cultivated in a 50 cm × 50 cm planting bag filled with PM measuring 20 cm in height and 30 cm in diameter. PM at the time of sowing has a pH of 6.9 ± 0.2 and soil moisture of 75%. Cultivation is carried out in an open space without any obstructions to sunlight. Watering is done in the morning and evening sufficiently on days without rain. NPK fertilization is carried out 7 days after planting (DAP) and 14 DAP, as much as 3 g/bag. EMMST or EAIST (50 cc) was applied to the respective samples at 14 and 28 DAP. Weeding is carried out if necessary.
2.6 Statistical analysis
Data were analyzed using SPSS v.22 and are presented as mean ± standard deviation of 10 replicates. A detailed description of all variables included in these analyses is provided in Table 2. The normality of data distribution was verified using the Shapiro-Wilk test, and the homogeneity of variances was confirmed with Levene’s test. For data meeting parametric assumptions, a one-way ANOVA was performed to determine significant differences among treatment means. Relationships between growth parameters were examined using correlation analysis. Model fitting was conducted to identify the best-fitting regression equations describing growth patterns over time. To evaluate interactions between treatment effects and physiological parameters, a Generalized Linear Model (GLM) was employed where appropriate. Model selection was based on goodness-of-fit statistics. For all analyses, differences were considered statistically significant at p < 0.05.
Table 2. Research variables and definitions
|
Variables (Code) |
Definition |
|
Fusarium sp. infection (FI) |
The diagnostic symptoms of infection consist of browning of vascular tissues, epinastic leaf curvature, growth retardation, gradual wilting, premature leaf drop, and eventual plant mortality [28]. |
|
Leaf height (LH) |
Average leaf height is measured from the base of the leaf to the tip of each clump (mm) |
|
Leaf diameter (LD) |
Average diameter of leaves per clump (mm) |
|
Leaf area (LA) |
Area of all leaves per clump (mm2) |
|
Leaf count (LC) |
The number of leaves per clump |
|
Tiler count (TC) |
Average number of bulbs per clump |
|
Root length (RL) |
The length of the root system from the base of the bulb to the tip of the longest root, measured using a ruler after carefully washing the roots free of planting media per clump |
|
Above-ground biomass (AGB) |
Average weight of all plant parts located above the soil surface (leaves and shoots), measured after harvesting per clump (mg) |
|
Below-ground biomass (BGB) |
Average weight of the entire root system, measured after carefully washing the roots free of planting media per clump (mg) |
|
Synergy Index (SI) |
An index quantifying the synergistic efficiency of biomass allocation toward leaf development. It was calculated using the formula: $\text{SI}=\frac{\text{ABG}}{(\text{ABG/LH)+(ABG/LC)}}$, where higher values indicate a more efficient conversion of biomass into vertical growth and leaf proliferation, reflecting a synergistic balance between biomass production and morphological development. |
3.1 Material transformation and physicochemical properties
The processing leaves from MM and AI into bioactive coatings reveals significant differences in their physicochemical properties. A key visual distinction observed during drying is that MM leaves change color from green to yellowish-brown, suggesting an increase in anthocyanins. In contrast, AI leaves maintain a relatively stable green pigmentation, indicating the retention of chlorophyll content. Furthermore, the aqueous extract of MM is more acidic (pH 6.2 ± 0.1) than that of AI (pH 6.8 ± 0.1). The most notable difference is in the final product’s rheology: the coating derived from MM forms a viscous gel, while the AI remains a thin slurry (Figure 2).
Table 3. Phytochemical compounds exhibiting plant growth regulatory effects
|
Melastoma malabathcirum |
Azadirachta indica |
||
|
Phytochemical Compounds |
Source/Reference |
Phytochemical Compounds |
Source/Reference |
|
Betulinic acid; Ellagic acid; Quercetin; Procyanidin B2; Gallic acid; Ursolic acid; Asiatic acid; Naringenin; Rutin; Malvidin-3-5-diglucoside; β-sitosterol; 1,4,6-Tri-O-galloyl-β-Dglucoside; Kaempferol; Kaempferol-3-O-β-Dglucoside; p-Hydroxybenzoic acid |
[29] |
Hyperoside; Nimbolide; Quercetin; Quercitrin; Rutin; β-Sitosterol; Nimbandiol |
[20] |
|
Epicatechin; Quercetin; Isoquercitrin; Citric acid; Quercitrin; Rutin; 7-hydroxycoumarine |
[30] |
Chlorogenic acid; luteolin-7-O-glucuronide |
[31] |
|
Arjunic acid; Ellagic acid; Maslinic acid; Corilagin; Neochlorogenic acid; Kaempferol; Quercetin; Astragalin; Rutin; Apigetrin; Asiatic acid; Tricoumaroyl spermidine; Quercitrin |
[32] |
Ascorbic acid; Cyclic trisulphide; tetrasulphide; 7-Hydroxyazadiradione |
[33] |
Figure 2. Sample material form
3.2 Phytochemical profile and bioactive potential
MM and AI produce diverse phytochemicals, including both species-specific compounds and a greater number of shared metabolites. Both plant species have been extensively utilized in traditional medicine, agriculture, and livestock management, with pharmacological studies validating the efficacy of their bioactive phytochemical constituents [33, 34]. While both species produce over 200 identified metabolites, Table 3 specifically highlights those with phytochemical compounds demonstrating plant growth regulatory activity. MM predominantly contains phenolic acids (e.g., gallic acid, ellagic acid) and flavonoids (e.g., quercetin), while AI is rich in terpenoids (e.g., nimbolide) and sulfur-containing compounds (e.g., trisulfide).
3.3 Biological efficacy: Disease suppression and plant growth
3.3.1 Disease onset and incidence
The initial onset of Fusarium wilt disease symptoms varied significantly among treatments. The control group exhibited the earliest signs of infection, with Fusarium symptoms first observed at 10 days after planting (DAP). In contrast, the PL, AI, and MM treatments showed delayed symptom appearance, with disease indications recorded at 12 DAP, 16 DAP, and 24 DAP, respectively. Notably, the MM treatment demonstrated the latest disease onset. The incidence of Fusarium wilt varied across treatments during the study period. Disease incidence was highest in the control group (4 cases), followed by the AI and PL treatments (3 cases each), and lowest in the MM treatment (2 cases). This indicates that the MM treatment has the greatest potential to inhibit Fusarium wilt.
3.3.2 Plant growth parameters
At 7 DAP, all bulbs remained healthy, exhibiting a green dot indicative of shoot potential. At 10 DAP, leaves displayed normal growth, and roots had developed, though shoots were absent in some bulbs. Differences among treatments became apparent at this stage. In the control group (C), 5 out of 10 bulbs developed shoots and leaves, with leaf height (LH) ranging from 5–75 mm and the leaf count (LC) between 1–4. The MM treatment showed the most advanced growth, with 9 out of 10 bulbs producing shoots and leaves. LH varied markedly (2-98 mm), and LC ranged from 1–7. In the AI group, 7 out of 10 bulbs had shoots and leaves, with LH measuring 3–40 mm and LC varying between 1–5. The PL treatment resulted in 8 out of 10 bulbs developing shoots and leaves, with LH and LC values comparable to the control (5–75 mm and 1–4, respectively). At 14 DAP, further divergence in growth and health was observed. One bulb (C9) decayed, while the others continued to grow, with green leaves increasing in height and count. All bulbs in the MM treatment thrived, exhibiting taller leaves and higher leaf counts. Notably, bulb MM10 hosted fungal colonisation (Figure 3); microscopic analysis revealed dark-green pigmentation, whorled phialides, and conidiophores with a tree-like structure, resembling Trichoderma sp.
Figure 3. Colonization of Trichoderma sp. on bulbs and growing media treated with M. malabathricum extract
3.4 Statistical analysis and synergistic relationship of growth
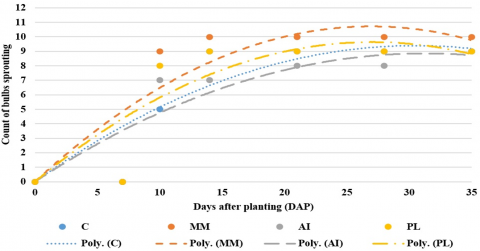
The cumulative count of bulbs showing a growth response throughout the study period is displayed in Figure 4. An outlier occurred in the AI treatment, where one bulb (AI1) produced a shoot at 32 DAP. Bulb Days to Sprouting (BDS) in all treatments followed quadratic polynomial relationships (Eq. (1)-Eq. (4)), where YBDS is the predicted BDS and DAP is days after planting.
Control:
YBDS = 0.615(DAP) - 0.010(DAP)2 (1)
R2 = 0.941; F = 39.950; p < 0.001.
MM:
YBDS = 0.795(DAP) - 0.015(DAP)2 (2)
R2 = 0.930; F = 33.391; p < 0.001.
AI:
YBDS = 0.566(DAP) - 0.009(DAP)2 (3)
R2 = 0.939; F = 38.332; p < 0.001.
PL:
YBDS = 0.713(DAP) - 0.013(DAP)2 (4)
R2 = 0.931; F = 33.840; p < 0.001.
Figure 4. Best-fit quadratic models of bulb days to sprouting
Analysis of first-order coefficients revealed that the MM treatment produced superior BDS to the control, with PL treatment showing intermediate performance and AI treatment demonstrating inferior results. The pattern implies that growth-stimulating phytochemicals may be present in MM, while growth-inhibiting compounds likely exist in AI. Partial correlations analysis identified significant positive relationships between leaf height and leaf count (r = 0.546; p < 0.001) and between above-ground biomass and below-ground biomass (r = 0.646; p < 0.001). These strong interdependencies suggest a synergistic effect may exist, where improvements in one growth parameter co-occur with enhancements in another, potentially indicating coordinated resource allocation or physiological coupling between plant organs.
In contrast to other treatments, the AI treatment exhibited statistically superior performance (Table 4), demonstrating 28–146% greater leaf area and 12–38% larger leaf diameter, whereas the PL treatment showed significantly longer roots (11–51% increase). While not statistically significant, the AI treatment demonstrated numerically superior values for leaf height, leaf count, above-ground biomass, and synergy index, while MM showed increased tiller count. Below-ground biomass was comparable across treatments, with MM exhibiting marginally lower values.
Table 4. ANOVA summary of shallot growth response variables at 35 DAP based on treatments
|
Growth Response |
Value (mean ± SD) |
Welch’s F (df1, df2) |
|||
|
C (N=8) |
MM (N=9) |
AI (N=9) |
PL (N=9) |
|
|
|
Root length (mm) |
91.75 ± 27.71 |
97.25 ± 21.82 |
123.89 ± 51.53 |
138.71 ± 31.96 |
3.618 (3, 18.245) * |
|
Leaf diameter (mm) |
2.04 ± 0.46 |
1.80 ± 0.18 |
2.50 ± 0.14 |
2.22 ± 0.37 |
26.235 (3, 16.893) ** |
|
Leaf area (mm2) |
603.80 ± 332.25 |
381.87 ± 117.75 |
941.00 ± 363.30 |
731.49 ± 297.37 |
8.656 (3, 17.528) ** |
|
Leaf height (avg., mm) |
166.51 ± 37.59 |
148.88 ± 37.25 |
186.14 ± 65.10 |
184.96 ± 40.43 |
1.443 (3, 19.672) |
|
Leaf count |
10.00 ± 2.93 |
11.56 ± 3.71 |
12.67 ± 4.61 |
10.56 ± 3.64 |
0.753 (3, 20.114) |
|
Tiller count |
2.63 ± 0.74 |
3.44 ± 0.88 |
3.11 ± 1.17 |
3.00 ± 0.50 |
1.355 (3, 18.956) |
|
Above-ground biomass (g) |
2.21 ± 1.31 |
2.21 ± 1.13 |
4.12 ± 2.49 |
3.27 ± 1.44 |
2.001 (3, 17.832) |
|
Below-ground biomass (g) |
1.21 ± 0.59 |
0.90 ± 0.33 |
1.21 ± 0.52 |
1.21 ± 0.70 |
1.064 (3, 19.447) |
|
Synergy Index |
9.94 ± 2.92 |
11.05 ± 3.71 |
12.59 ± 4.62 |
10.94 ± 3.90 |
0.637 (3, 20.338) |
Table 5. Generalized linear models summary
|
Parameter |
B |
Std. Error |
95% Wald CI |
Hypothesis Test |
|||
|
Lower |
Upper |
Wald Chi-Square |
df |
Sig. |
|||
|
(Intercept) |
2.569 |
4.57 |
-6.381 |
11.520 |
0.317 |
1 |
0.574 |
|
RL |
0.032 |
.01 |
0.010 |
0.054 |
8.287 |
1 |
0.004 |
|
LD |
2.553 |
2.94 |
-3.204 |
8.310 |
0.755 |
1 |
0.385 |
|
LA |
-0.001 |
.005 |
-0.011 |
0.008 |
0.080 |
1 |
0.777 |
|
T_PL |
-7.069 |
2.78 |
-12.509 |
-1.629 |
6.487 |
1 |
0.011 |
|
T_AI |
-8.547 |
3.60 |
-15.604 |
-1.490 |
5.635 |
1 |
0.018 |
|
T_MM |
-7.671 |
3.37 |
-14.272 |
-1.069 |
5.186 |
1 |
0.023 |
|
T_C |
0a |
. |
. |
. |
. |
. |
. |
|
T_PL * LA |
0.008 |
.004 |
0.001 |
0.016 |
4.478 |
1 |
0.034 |
|
T_AI * LA |
0.010 |
.004 |
0.001 |
0.019 |
5.140 |
1 |
0.023 |
|
T_MM * LA |
0.023 |
.006 |
0.010 |
0.035 |
12.831 |
1 |
0.000 |
|
T_C * LA |
0a |
. |
. |
. |
. |
. |
. |
|
(Scale) |
4.879b |
1.22 |
2.989 |
7.965 |
|
|
|
The generalized linear model analysis revealed significant treatment-specific effects on the synergy index (SI), demonstrating how physiological parameters interact to influence overall plant performance (Table 5). Treatments exerted statistically significant main effects (Wald χ² = 8.567, p = 0.036), with particularly strong treatment × leaf area interactions (Wald χ² = 13.811, p = 0.003). Notably, the MM treatment showed the most pronounced synergistic relationship with leaf area (β = 0.023, p < 0.001), suggesting that its growth-promoting effects become substantially more effective as foliar expansion increases. This pattern was similarly observed, though to lesser degrees, in AI (β = 0.010, p = 0.023) and PL (β = 0.008, p = 0.034) treatments. While all treatments exhibited negative direct effects on SI compared to controls (AI: β = -8.547; MM: β = -7.671; PL: β = -7.069; all p < 0.05), these were counterbalanced by their positive interactions with leaf area. Root length emerged as an independent positive predictor of SI (β = 0.032, p = 0.004), underscoring its role in growth synergy, whereas leaf diameter showed no significant association (p = 0.385).
These findings collectively suggest that treatment efficacy is modulated by leaf development status, with MM displaying the strongest capacity to convert foliar expansion into whole-plant growth synergy. The dual significance of root elongation and treatment-specific leaf area responses highlights the integrated nature of plant growth regulation, where above- and below-ground processes interact to determine overall performance.
4.1 Linking material properties to functionality
The distinct physicochemical properties of the MM and AI extracts directly inform their functionality as bioactive coatings. The color change in MM leaves from green to yellowish-brown during drying suggests an increase anthocyanin, a group of unstable acidic compounds comprising at least 71 phytochemical compounds divided into 8 subgroups: cyanidin (16), peonidin (11), delphinidin (10), pelargonidin (9), flavonoids (8), malvidin (7), petunidin (6) and procyanidins (4) [35]. Conversely, the stable green pigmentation of AI leaves indicates successful chlorophyll retention, aligning with studies showing high phytochemical preservation at low drying temperature [36, 37]. The most notable difference was the rheology: MM formed a viscous gel while AI remained a thin slurry. This is likely due to the interaction of anthocyanins with starch, which can alter the material’s microstructure and chemical-physical properties [38, 39]. These properties necessitate advanced stabilization strategies for liquid applications, with cassava starch emerging as an optimal coating matrix due to its low amylose content and high viscosity [40]. While its neutral characteristics enable versatile applications, coating performance critically depends on additive selection – as evidenced by the trade-off between sorbitol’s superior water-vapor permeability but inferior antioxidant preservation compared to glycerol [41, 42].
Research findings indicating increased antioxidant compounds in dried MM and AI leaves are supported by prior studies showing that drying technologies not only facilitate phytochemical handling but also enhance bioactive content, such as phenolic and alkaloids, offering a sustainable solution for plant-based applications [43]. While dried materials offer straightforward handling, maintaining phytochemical efficacy in liquid extracts necessitates careful degradation prevention and storage optimization. This is particularly relevant for MM and AI extracts, which demonstrate marked sensitivity to environmental factors – showing rapid degradation under oxygen exposures [44], elevated temperatures/alkaline pH [45], and UV/microbial activity [46]. These stability limitations underscore the need for advanced stabilization approaches in phytochemical applications.
4.2 Mechanisms of biological action
The observed biological efficacy–delayed disease onset and enhanced plant growth–can be directly correlated to the phytochemical profile of each species. The MM treatment demonstrated the latest onset of Fusarium wilt and the lowest disease incidence. This superior antifungal activity is likely multifactorial. Anthocyanin-rich extracts are known to inhibit Fusarium conidial germination [47], while specific compounds like rutin found in MM act as alpha-glucosidase inhibitors and disrupt Fusarium effector proteins (SIXPs) and biofilms [30, 48]. Furthermore, the unique colonization by Trichoderma sp. on an MM-treated bulb (Figure 3) suggests a synergistic relationship [49]. Furthermore, the efficacy against Fusarium is supported by the documented mode of action of phytochemicals. Phytochemicals exhibit broad-spectrum antifungal activity through multiple synergistic mechanisms, including cell wall disruption, plasma membrane damage, genetic material interference, biofilm inhibition, and hyphal growth suppression [50].
The application of phenolic compounds, including quercetin, has also been reported to stimulate root expansion [51], a finding consistent with our results. The quercetin-induced increase in chlorophyll levels not only mitigates iron chlorosis but also directly enhances plant biomass accumulation through improved photosynthetic efficiency [52]. Coupled with its growth-promoting capacity (up to 117% in ryegrass [53]) and pathogen-inhibiting properties [54], quercetin demonstrates triple-action potential in crop management. Conversely, certain phenolic acids present in AI leaves–namely ferulic, caffeic, and vanillic acid [55]–are known for their allelopathic effects [51], which account for the relative delay in bulb sprouting observed in the AI treatment. Beyond direct antifungal and plant growth-promoting action, AI leaf extract effectively counteracts oxidative damage induced by water stress, leading to improved overall plant growth [56]. Furthermore, phytochemical compounds derived from MM ameliorate acidic soils by directly reducing available Fe and exchangeable Al, and indirectly promoting the availability of P, N, and K [57] through metal chelation and stimulation of microbial activity, thereby enhancing soil microbial diversity, which is beneficial for plant health.
Our results reveal that MM treatment uniquely enhances both plant performance (bulb sprouting, Fusarium resistance) and beneficial Trichoderma colonization, contrasting with AI treatment effects. This dual functionality aligns with established mechanisms whereby: (1) Trichoderma coatings promote plant growth [58], and (2) Trichoderma seed pretreatment upregulates anthocyanin biosynthesis [59]. Beyond merely documenting the efficacy of the coatings, our study proposes a novel, self-reinforcing mechanistic model (Figure 5) that integrates direct phytochemical action with beneficial microbiome recruitment. This synergistic feedback loop, particularly prominent in the MM treatment, explains the sustained resistance and growth promotion observed. Furthermore, the model elucidates the trade-off between direct antifungal strength and growth inhibitory effects in AI, providing a nuanced understanding of plant-phytochemical-pathogen interactions.
Figure 5. Proposed self-reinforcing mechanism of M. malabathricum bioactive coating for enhanced shallot resilience on peatland
These findings expand the documented benefits of anthocyanin coatings in seed preservation [60], while highlighting formulation constraints – excessive concentrations may compromise tensile strength and water-vapor permeability despite improved phytochemical stability [61]. While our experimental trials demonstrated the coating’s multifunctional efficacy–enhancing bulb sprouting, Fusarium resistance, and Trichoderma colonization–scalable adoption requires addressing socio-economic constraints, such as raw material accessibility and cost-efficiency compared to conventional treatments.
While this study demonstrates the clear superiority of MM over AI in promoting synergistic growth and Trichoderma recruitment, we acknowledge that the comparative analysis primarily highlighted their mechanistic differences rather than a comprehensive efficacy ranking. The distinct performance -where MM excelled in growth synergy and microbiome recruitment, while AI showed stronger direct antifungal properties but allelopathic effects- reveals a fundamental trade-off between different modes of action. This functional contrast underscores that “effectiveness’ is multi-dimensional, and the optimal choice of plant extract may depend on specific agricultural priorities: growth promotion and soil health (favoring MM) versus immediate pathogen suppression (where AI has value). Future formulations could even explore combining these extracts to leverage their complementary strengths.
4.3 Socio-economic and environmental perspectives
The International Union for Conservation of Nature (IUCN) classifies AI as a least concern species with a stable population across South and Southeast Asia [62], while MM remains unassessed. However, studies indicate MM is invasive [63] and adapts well to degraded soil [64], suggesting both species are viable bioresources with low conservation risks. Consequently, their availability is predicted to be abundant, making them economically affordable or even free. In the Indonesian context, tapioca flour holds a distinct economic and practical advantage over other starches. This economic resilience stems from its supply chain independence; unlike imported wheat flour, cassava is cultivated domestically, creating a more stable value shield from foreign exchange volatility. This is complemented by its ready market availability and simple processing technology [65].
This study establishes MM as a safer alternative to synthetic fungicides, addressing ecological and health risks from persistent residues [66, 67]. Quantitative analysis reveals a nuanced efficacy profile: while the chlorothalonil treatment showed superior root development (138.71 ± 31.96 mm), the MM coating demonstrated comparable Fusarium wilt suppression with the added advantage of significantly enhanced growth synergy. This dual functionality -direct pathogen suppression coupled with microbiome recruitment and growth promotion-represents a key advantage over single-mode chlorothalonil. For farmer adoption, bridging the yield gap with conventional chemical remains crucial; however, the multifunctional benefits of MM may offer compelling value where soil health and sustainable practices are prioritized.
4.4 Statistical robustness and study limitations
The statistical analysis provides robust evidence for the treatment-specific effects, demonstrated by the highly significant (p < 0.001) quadratic models of BDS (R2 ≥ 0.930) and the compelling treatment × leaf area interaction in the GLM (Wald χ² = 13.811, p = 0.003). These results, coupled with large effect sizes (Cohen’s f = 0.55-1.48) for key growth parameters, strongly confirm that the MM coating dynamically enhances growth synergy in a leaf development-dependent manner, despite the sample size (N = 40) and 35-day duration being optimized for proof-of-concept rather than long-term yield assessment.
However, certain limitations contextualize these findings. The proposed mechanism involving Trichoderma colonization, while observationally supported, lacks molecular validation. Furthermore, the reliance on natural pathogen pressure–though ecologically relevant–precludes precise pathogen identification, and the exclusion of coating mechanical properties represents a necessary trade-off between practical applicability and material standardization. These limitations do not diminish the statistical robustness of our central findings but precisely define critical pathways for future research, particularly regarding microbial dynamics, molecular characterization, and extraction standardization for commercial scaling.
This study demonstrates the profound potential of MM and AI leaf extracts as bioactive coatings for sustainable shallot cultivation. The distinct physicochemical properties of each extract—a viscous, acidic gel for MM and a neutral, thin slurry for AI—directly stem from their unique phytochemical compositions, rich in flavonoids/phenolic acids and terpenoids/sulfur compounds, respectively. Biological efficacy assays revealed a critical trade-off: while both coatings delayed Fusarium wilt onset, the MM coating provided superior, synergistic growth promotion and the highest level of disease suppression. This enhanced performance is attributed to a proposed self-reinforcing mechanism where MM phytochemicals not only act directly against the pathogen but also facilitate beneficial Trichoderma colonization, which in turn bolsters host defenses. In contrast, the AI coating's allelopathic properties, though antifungal, impeded early growth, resulting in inferior performance.
Statistical modeling quantitatively confirmed MM's superiority, showing it had the highest growth coefficient and the most significant synergistic interaction between treatment application and leaf area development. These findings collectively position the MM-based coating as a multifaceted, sustainable technology that concurrently addresses disease management and growth enhancement. Therefore, this research validates a novel plant-based strategy to reduce dependency on synthetic fungicides, paving the way for more resilient agricultural systems. The next phase of research should focus on stabilizing anthocyanin–starch interactions within the coating matrix, preventing oxidation and pH-induced degradation that limit field durability. Equally important is quantifying Trichoderma recruitment dynamics and phytochemical persistence across soil moisture gradients in peatland environments to validate the long-term efficacy of this mechanism.
We acknowledge to Regional Research and Development Agency of South Sumatra Province, Indonesia, for funding and all who have contributed to this research.
[1] van Oort, P.A.J., Timmermans, B.G.H., Schils, R.L.M., Van Eekeren, N. (2023). Recent weather extremes and their impact on crop yields of the Netherlands. European Journal Agronomy, 142: 126662. https://doi.org/10.1016/j.eja.2022.126662
[2] Winona, B., Sawitri, W.D., Idris, M., Handayani, V.D.S., Wibowo, A., Matra, D.D., Doni, F., Widiastuti, A. (2025). Dynamics of defense-related genes expression in shallot after UV-B-induced resistance against Fusarium acutatum showed time difference. Physiological and Molecular Plant Pathology, 136: 102559. https://doi.org/10.1016/j.pmpp.2024.102559
[3] Central Statistics Agency (BPS - Statistics Indonesia). (2025). Annual Fruit and Vegetable Crop Production by Crop Type, 2024. https://www.bps.go.id/id/statistics-table/3/WXpSVU5uUTBOSEl5WVhGQmVESTVSVnBSVlhWeVVUMDkjMyMwMDAw/produksi-tanaman-buah-buahan-dan-sayuran-tahunan-menurut-jenis-tanaman.html?year=2024.
[4] Central Statistics Agency (BPS - Statistics Indonesia). (2025). Harvested Area of Annual Vegetable and Fruit Crops by Crop Type, 2024. https://www.bps.go.id/id/statistics-table/3/VTNWM01VdGhTelZTTTNSS1NFSkVZazkzWjJKcWR6MDkjMyMwMDAw/luas-panen-tanaman-sayuran-dan-buah-buahan-semusim-menurut-jenis-tanaman.html?year=2024.
[5] Ismayaningrum, D., Prasetya, R., Harris, I., Ilyasa, R.L., Romdhan, A., Warsini. (2025). Performance report of the directorate general of Horticulture. Ministry of Agriculture of the Republic of Indonesia. https://hortikultura.pertanian.go.id/laporan-kinerja-direktorat-jenderal-hortikultura/.
[6] Suravi, T.I., Hasan, M.K., Jahan, I., Shopan, J., Saha, M., Debnath, B., Ahammed, G.J. (2024). An update on post-harvest losses of onion and employed strategies for remedy. Scientia Horticulturae, 338: 113794. https://doi.org/10.1016/j.scienta.2024.113794
[7] Sahara, D., Yaumidin, U.K., Suhendrata, T., Setiani, C., Beti, J.A., Dewi, T., Fadwiwati, A.Y., Idaryani, Atman, Yardha, Waas, E., Asnawi, R., Syam, A. (2025). Sustainability of shallot farming system in lowland Central Java Province, Indonesia: MICMAC analysis approach. Environmental Challenges, 20: 101212. https://doi.org/10.1016/j.envc.2025.101212
[8] da Silva Pereira, L., Salomón, A.L., Bermudez, V.A.C., Rius, S.P. (2025). Bioactive proteins from Trichoderma for the control of Fusarium: Evaluation of antagonist efficacy and potential for agronomic application. Biocatalysis and Agricultural Biotechnology, 67: 103628. https://doi.org/10.1016/j.bcab.2025.103628
[9] Shi, Z., Zhang, M., Chen, G., Cao, A., Wang, Q., Yan, D., Fang, W., Li, Y. (2025). Evaluation of newly combination of Trichoderma with dimethyl disulfide fumigant to control Fusarium oxysporum, optimize soil microbial diversity and improve tomato yield. Ecotoxicology and Environmental Safety, 292: 117903. https://doi.org/10.1016/j.ecoenv.2025.117903
[10] Rahman, M.M., Sultana, N., Hoque, M.A., Azam, M.G., Islam, M.R., Hossain, M.A. (2024). Conservation tillage (CT) for climate-smart sustainable intensification: Benchmarking CT to improve soil properties, water footprint and bulb yield productivity in onion cultivation. Heliyon, 10(22): e39749. https://doi.org/10.1016/j.heliyon.2024.e39749
[11] Shelake, P., Mohapatra, D., Sabat, M. (2025). Impact of gaseous ozone treatment on storability and quality of onion (Allium cepa L.) during bulk storage. Journal of Stored Products Research, 112: 102656. https://doi.org/10.1016/j.jspr.2025.102656
[12] Poursakhi, S.R., Asadi-Gharneh, H.A., Nasr-Esfahani, M., Abbasi, Z., Khankahdani, H.H. (2025). Defense-related enzymes associated with resistance to onion Fusarium basal rot. Plant Physiology and Biochemistry, 219: 109326. https://doi.org/10.1016/j.plaphy.2024.109326
[13] Sakane, K., Akiyama, M., Ando, A., Shigyo, M., Ito, S., Sasaki, K. (2023). Identification of a novel effector gene and its functional tradeoff in Fusarium oxysporum f. sp. cepae that infects Welsh onion. Physiological and Molecular Plant Pathology, 123: 101939. https://doi.org/10.1016/j.pmpp.2022.101939
[14] Poursakhi, S., Asadi-Gharneh, H.A., Nasr-Esfahani, M., Abbasi, Z., Khankahdani, H.H. (2024). Identification of novel associations of candidate marker genes with resistance to onion-fusarium basal rot interaction pathosystem. Plant Gene, 37: 100440. https://doi.org/10.1016/j.plgene.2023.100440
[15] Gökçe, A.F., Gökçe, Z.N.Ö., Junaid, M.D., Chaudhry, U.K. (2023). Evaluation of biochemical and molecular response of onion breeding lines to drought and salt stresses. Scientia Horticulturae, 311: 111802. https://doi.org/10.1016/j.scienta.2022.111802
[16] Kauffman, J.B., Adame, M.F., Adinugroho, W.C., Anshari, G., et al. (2025). Total ecosystem carbon stocks of tropical peat forests and greenhouse gas emissions from their disturbance. Forest Ecology and Management, 593: 122840. https://doi.org/10.1016/j.foreco.2025.122840
[17] Pemerintah Republik Indonesia. (2016). Government Regulation (PP) No. 57 of 2016: Amendment to Government Regulation Number 71 of 2014 concerning the Protection and Management of Peat Ecosystems. https://peraturan.bpk.go.id/Details/5778/pp-no-57-tahun-2016.
[18] Pittroff, S.M., Brems, A.R., Brunshøj, R.J., Christiansen, J.V., et al. (2024). Novel rapid screening assay to incorporate complexity and increase throughput in early-stage plant biological testing. Rhizosphere, 30: 100897. https:/doi.org/10.1016/j.rhisph.2024.100897
[19] Stasiun Klimatologi Sumatera Selatan. Climate Parameter Analysis. https://staklim-sumsel.bmkg.go.id/analisis-parameter-iklim.
[20] US Department of Agriculture. Dr. Duke’s Phytochemical and Ethnobotanical Databases. https://phytochem.nal.usda.gov/.
[21] Kim, S., Chen, J., Cheng, T., Gindulyte, A., et al. (2025). PubChem 2025 update. Nucleic Acids Research, 53(D1): D1516-D1525. https://doi.org/10.1093/nar/gkae1059
[22] Eskandarzade, P., Mehrjerdi, M.Z., Gruda, N.S., Aliniaeifard, S. (2024). Phytochemical compositions and antioxidant activity of green and purple basils altered by light intensity and harvesting time. Heliyon, 10(10): e30931. https://doi.org/10.1016/j.heliyon.2024.e30931
[23] Qin, Y., Xu, F., Yuan, L., Hu, H., Yao, X., Liu, J. (2020). Comparison of the physical and functional properties of starch/polyvinyl alcohol films containing anthocyanins and/or betacyanins. International Journal of Biological Macromolecules, 163: 898-909. https://doi.org/10.1016/j.ijbiomac.2020.07.065
[24] Kim, J., Lim, H., Jang, Y., Park, M., Moon, D.H., Lee, G., Hyun, S. (2025). Effect of drying treatment on the leachability of metallic elements (Zn, As, Cd, and Pb) from amended mine soils during batch leaching experiments. Environmental Pollution, 374: 126246. https://doi.org/10.1016/j.envpol.2025.126246
[25] Xiao, D., Tang, X., Chen, S., Chu, G., Liu, Y., Wang, D., Xu, C. (2025). Aeration treatment promotes transformation of soil phosphorus fractions to plant-available phosphorus by modulating rice rhizosphere microbiota. Soil Tillage Research, 245: 106318. https://doi.org/10.1016/j.still.2024.106318
[26] Suwandi, Sumarni, N., Setiawati, W. (2013). Off-season shallot cultivation technology. Badan Penelitian dan Pengembangan Pertanian Kementerian Pertanian, Jakarta, Indonesia. https://ppid.pertanian.go.id/doc/1/budidaya%20bawang%20merah%20di%20luar%20musim.pdf.
[27] Hermanto, C., Maharijaya, A., Arsanti, I.W., Hayati, M., et al. (2017). Guidelines for cultivating red onions using seeds. Directorate of Vegetables and Medicinal Plants, Ministry of Agriculture, Jakarta, Indonesia. https://hortikultura.pertanian.go.id/wp-content/uploads/2021/03/PEDOMAN-BUDIDAYA-BAMER-2017.pdf.
[28] Sampaio, A.M., de Sousa Araújo, S., Rubiales, D., Vaz Patto, M.C. (2020). Fusarium wilt management in legume crops. Agronomy, 10(8): 1073. https://doi.org/10.3390/agronomy10081073
[29] Kumar, V., Sachan, R., Rahman, M., Rub, R.A., et al. (2021). Chemopreventive effects of Melastoma malabathricum L. extract in mammary tumor model via inhibition of oxidative stress and inflammatory cytokines. Biomedicine & Pharmacotherapy, 137: 111298. https://doi.org/10.1016/j.biopha.2021.111298
[30] Lestari, O.A., Palupi, N.S., Setiyono, A., Kusnandar, F., Yuliana, N.D. (2024). LC-MS metabolomics and molecular docking approaches to identify antihyperglycemic and antioxidant compounds from Melastoma malabathricum L. Leaf. Saudi Journal of Biological Sciences, 31(8): 104047. https://doi.org/10.1016/j.sjbs.2024.104047
[31] Abdalla, M., Mohapatra, R.K., Sarangi, A.K., Mohapatra, P.K., et al. (2021). In silico studies on phytochemicals to combat the emerging COVID-19 infection. Journal of Saudi Chemical Society, 25(12): 101367. https://doi.org/10.1016/j.jscs.2021.101367
[32] Lee, T.H., Lee, C.H., Ong, P.Y., Wong, S.L., et al. (2022). Comparison of extraction methods of phytochemical compounds from white flower variety of Melastoma malabathricum. South African Journal of Botany, 148: 170-179. https://doi.org/10.1016/j.sajb.2022.04.026
[33] Dhakad, A.K., Kumar, R., Choudhary, R., Singh, S., Khan, S., Poonia, P.K. (2025). Traditional to modern perspectives on Neem (Azadirachta indica): A gateway to bioactive compounds, sustainable agrochemicals and industrial applications. Industrial Crops and Products, 231: 121155. https://doi.org/10.1016/j.indcrop.2025.121155
[34] Tiwari, M., Barooah, M.S., Bhuyan, D. (2023). Phytochemical and bioactive potentialities of Melastoma malabathricum. In Recent Frontiers of Phytochemicals, pp. 601-615. https://doi.org/10.1016/B978-0-443-19143-5.00024-4
[35] Fan, H., Liao, H., Shen, Y., Sani, M.N.H., Yong, J.W.H., Song, J. (2024). Unravelling the physiological and molecular mechanisms of leaf color change in Acer griseum through multi-omics analysis. Plant Physiology and Biochemistry, 216: 109198. https://doi.org/10.1016/j.plaphy.2024.109198
[36] Sinha, R., Thakur, A.D., Raj, R. (2024). Investigating drying behavior and quality of Neem leaves using a novel biomass gasification-powered climate control unit with built-in humidity control. International Communications in Heat and Mass Transfer, 158: 107888. https://doi.org/10.1016/j.icheatmasstransfer.2024.107888
[37] Nisha, Sharma, N., Mohite, A.M. (2025). Effect of vacuum and through flow drying technique on mathematical modelling, functional properties, color degradation kinetics, and essential oil components of fish mint (Houttuynia cordata Thunb). Food Physics, 2: 100057. https://doi.org/10.1016/j.foodp.2025.100057
[38] Dangles, O., Fenger, J.A. (2018). The chemical reactivity of anthocyanins and its consequences in food science and nutrition. Molecules, 23(8): 1970. https://doi.org/10.3390/molecules23081970
[39] Ou, S.J.L., Yu, J., Zhou, W., Liu, M.H. (2022). Effects of anthocyanins on bread microstructure, and their combined impact on starch digestibility. Food Chemistry, 374: 131744. https://doi.org/10.1016/j.foodchem.2021.131744
[40] Das, S., Das, M.K., Jamatia, T., Bhattacharya, B., et al. (2024). Advances of cassava starch-based composites in novel and conventional drug delivery systems: A state-of-the-art review. RSC Pharmaceutics, 1(2): 182-203. https://doi.org/10.1039/D3PM00008G
[41] Sivapragasam, N., Maqsood, S., Rupasinghe, H.P.V. (2024). Berry bioactive compounds immobilized in starch matrix for active and intelligent packaging: A review. Future Foods, 10: 100397. https://doi.org/10.1016/j.fufo.2024.100397
[42] Costa, B.P., Carpine, D., Ikeda, M., Pazzini, I.A.E., da Silva Bambirra Alves, F.E., de Melo, A.M., Ribani, R.H. (2023). Bioactive coatings from non-conventional loquat (Eriobotrya japonica Lindl.) seed starch to extend strawberries shelf-life: An antioxidant packaging. Progress in Organic Coatings, 175: 107320. https://doi.org/10.1016/j.porgcoat.2022.107320
[43] Zhao, Y., Zhang, W., Yang, H., Xu, Z., Wang, X., Zhang, Z., Deng, J. (2025). Effects of drying methods on phytochemicals and antioxidant activity of broccoli by-products. Food Research International, 208: 116284. https://doi.org/10.1016/j.foodres.2025.116284
[44] Shehata, E., Percival, J., Philo, M., Needs, P.W., Clifford, M.N., Kroon, P.A. (2025). Aerobic in vitro studies of spontaneous anthocyanin degradation are not useful models for anthocyanin breakdown in the human colon. Food Chemistry, 486: 144614. https://doi.org/10.1016/j.foodchem.2025.144614
[45] Idir, S., Achat, S., Cruz, L., Dangles, O. (2025). Anthocyanin-rich extracts: Susceptibility to color loss by hydration and thermal degradation, influence of metal ions and endogenous copigments. Food Chemistry, 481: 144004. https://doi.org/10.1016/j.foodchem.2025.144004
[46] Riyajan, S.A., Chantawee, K. (2020). Cassava starch composite based films for encapsulated neem: Effect of carboxylated styrene-butadiene rubber coating. Food Packaging and Shelf Life, 23: 100438. https://doi.org/10.1016/j.fpsl.2019.100438
[47] Felici, L., Atanasova, V., Ponts, N., Ducos, C., Francesconi, S., Sestili, F., Richard-forget, F., Balestra, G.M. (2025). Cyanidin 3-O-glucoside and other anthocyanins affect enniatins production in Fusarium avenaceum. Fungal Bioogy, 129(6): 101640. https://doi.org/10.1016/j.funbio.2025.101640
[48] Li, Y., Tian, X., Zhang, L., Lin, J., et al. (2025). Rutin resists Aspergillus fumigatus keratitis by activating Nrf2/HO-1 pathway, inhibiting Dectin-1/p-Syk pathway and affecting fungal structures. Experimental Eye Research, 254: 110323. https://doi.org/10.1016/j.exer.2025.110323
[49] Cournault, Q., Gibot-Leclerc, S., Monnier, N., Steinberg, C. (2025). Modes of action and inhibitory activity of Trichoderma species on potato and tomato pathogenic Phytophthora infestans: A review. Fungal Biology Reviews, 52: 100414. https://doi.org/10.1016/j.fbr.2025.100414
[50] da Silva, V.B., Almeida-Bezerra, J.W., Novais, M.H.G., Farias, N.S., et al. (2024). Chemical composition, antifungal, and anti-virulence action of the stem bark of Hancornia speciosa Gomes (Apocynaceae) against Candida spp. Journal of Ethnopharmacology, 321: 117506. https://doi.org/10.1016/j.jep.2023.117506
[51] Hurtado-Navarro, M., Garcia-Ibañez, P., Pascual, J.A., Carvajal, M. (2025). Interaction of beneficial microorganisms and phenolic compounds in hydroponically cultivated tomato plants. Plant Physiology and Biochemistry, 222: 109793. https://doi.org/10.1016/j.plaphy.2025.109793
[52] Pestana, M., Saavedra, T., Gama, F., Rodrigues, M.A., de Varennes, A., Da Silva, J.P., Correia, P.J. (2024). Quercetin promotes the recovery of iron chlorosis in strawberry plants. Plant Physiology and Biochemistry, 217: 109266. https://doi.org/10.1016/j.plaphy.2024.109266
[53] Toshmatov, Z., Zhou, S., Huang, L., Kuchkarova, N., et al. (2025). Flavonoids of Solanum rostratum seeds: Identification and plant growth regulatory activity evaluation. Natural Product Research, 1-6. https://doi.org/10.1080/14786419.2025.2526733
[54] Shadidizaji, A., Cinisli, K.T., Warda, M., Cicek, B., Hacimuftoglu, A. (2024). Virtual insights into the quercetin-Melampsora lini-derived effector AvrM14 interaction: An In silico exploration of plant defense mechanisms. Physiological and Molecular Plant Pathology, 129: 102200. https://doi.org/10.1016/j.pmpp.2023.102200
[55] Nazir, F., Jabeen, Z., Aslam, F., Mohammed, O.A., Ahmad, N., Igbal, S., Nazir, A., Iqbal, M. (2025). Unveiling multifaceted bioactivity assessment of Psidium guajava and Azadirachta indica leaves extract as a potential natural tyrosinase inhibitors. Biocatalysis and Agricultural Biotechnology, 64: 103486. https://doi.org/10.1016/j.bcab.2024.103486
[56] Naz, H., Akram, N.A., Ashraf, M., Ingo Hefft, D., Latief Jan, B. (2022). Leaf extract of neem (Azadirachta indica) alleviates adverse effects of drought in quinoa (Chenopodium quinoa Willd.) plants through alterations in biochemical attributes and antioxidants. Saudi Journal of Biological Sciences, 29(3): 1367-1374. https://doi.org/10.1016/j.sjbs.2022.01.038
[57] Sulaiman, S., Liew, K. (2025). Evaluating the effects of Melastoma malabathricum and Dicranopteris linearis as amendments for acid sulfate soil. Soil Advances, 3: 100035. https://doi.org/10.1016/j.soilad.2025.100035
[58] Younes, N.A., Anik, T.R., Rahman, M.M., Wardany, A.A., Dawood, M.F.A., Tran, L.P., Abdel Latef, A.A.H., Mostofa, M.G. (2023). Effects of microbial biostimulants (Trichoderma album and Bacillus megaterium) on growth, quality attributes, and yield of onion under field conditions. Heliyon, 9(3): e14203. https://doi.org/10.1016/j.heliyon.2023.e14203
[59] Elkelish, A.A., Alhaithloul, H.A.S., Qari, S.H., Soliman, M.H., Hasanuzzaman, M. (2020). Pretreatment with Trichoderma harzianum alleviates waterlogging-induced growth alterations in tomato seedlings by modulating physiological, biochemical, and molecular mechanisms. Environmental and Experimental Botany, 171: 103946. https://doi.org/10.1016/j.envexpbot.2019.103946
[60] Gao, L., Sun, H., Nagassa, M., Li, X., Pei, H., Liu, S., Gu, Y., He, S. (2024). Edible film preparation by anthocyanin extract addition into acetylated cassava starch/sodium carboxymethyl cellulose matrix for oxidation inhibition of pumpkin seeds. International Journal of Biological Macromolecules, 267: 131439. https://doi.org/10.1016/j.ijbiomac.2024.131439
[61] Zhu, F. (2025). Starch based films and coatings for food packaging: Interactions with phenolic compounds. Food Research International, 204: 115758. https://doi.org/10.1016/j.foodres.2025.115758
[62] Barstow, M., Deepu, S. (2018). Azadirachta indica. The IUCN Red List of Threatened Species. https://www.iucnredlist.org/species/61793521/61793525.
[63] Cheek, M.D., Boon, R.G.C., Wong, K.M., Hadebe, M. (2021). Assessment of the degree of establishment of five introduced Melastomataceae taxa in South Africa. South African Journal of Botany, 142: 502-508. https://doi.org/10.1016/j.sajb.2021.07.031
[64] Blackham, G.V., Webb, E.L., Corlett, R.T. (2014). Natural regeneration in a degraded tropical peatland, Central Kalimantan, Indonesia: Implications for forest restoration. Forest Ecology and Management, 324: 8-15. https://doi.org/10.1016/j.foreco.2014.03.041
[65] Rozi, F., Santoso, A.B., Mahendri, I.G.A.P, Hutapea, R.T.P., Wamaer, D., Siagian, V., Elisabeth, D.A., Sugiono, S., Handoko, H., Subagio, H., Syam, A. (2023). Indonesian market demand patterns for food commodity sources of carbohydrates in facing the global food crisis. Heliyon, 9(6): e16809. https://doi.org/10.1016/j.heliyon.2023.e16809
[66] Malhat, F., Saber, E.S., Shokr, S.A.S., Eissa, F. (2025). Pesticide residues in Egyptian vegetables: A comprehensive analysis of compliance, co-occurrence of multiple residues, and health risk assessment. Journal of Food Composition and Analysis, 143: 107634. https://doi.org/10.1016/j.jfca.2025.107634
[67] Wang, T., Wang, Z., Liao, G., Li, X., Gu, J., Qiu, J., Qian, Y. (2025). Carbendazim led to neurological abnormalities by interfering metabolic profiles in zebrafish brain after short-term exposure. Environmental Chemistry and Ecotoxicology, 7: 164-173. https://doi.org/10.1016/j.enceco.2024.12.001