Agusdin Dharma Fefirenta*![]() | I Putu Gede P. Damayanto
| I Putu Gede P. Damayanto![]() | Kusuma Rahmawati
| Kusuma Rahmawati![]() | Bayu Arief Pratama
| Bayu Arief Pratama![]() | Jalma Giring Sukmawati
| Jalma Giring Sukmawati![]()
© 2025 The authors. This article is published by IIETA and is licensed under the CC BY 4.0 license (http://creativecommons.org/licenses/by/4.0/).
OPEN ACCESS
Arbuscular mycorrhizal (AM) fungi are key components of terrestrial ecosystems, supporting plant nutrition, soil stability, and overall ecosystem function. However, their community structure within the unique biogeographical zone of Wallacea remains a critical knowledge gap. To address this, we characterized AM fungal communities along an elevational gradient (300–800 masl) in Kokolomboi Biodiversity Park, Banggai Islands, Central Sulawesi, Indonesia. Nine species from four genera were identified, with a notable dominance of Glomus boreale, which was associated with 76% of surveyed plants, suggesting a potential keystone role in these communities. Diversity values ranged from low to moderate (Shannon index 0.00–1.26), while evenness approached 1, suggesting balanced communities despite the low overall richness. This pattern may reflect strong environmental filtering shaped by the park’s unique geology. Crucially, only Acaulospora scrobiculata and Glomus multicaule showed significant correlations with available phosphorus (r = 0.87–0.94), identifying them as potential bioindicators of soil fertility. The results indicate that vegetation structure and soil chemistry are codependent drivers of AM fungal communities, with phosphorus availability emerging as the primary limiting factor.
Arbuscular mycorrhizal fungi, correlation, Shannon diversity index, Banggai Kepulauan, Wallacea, Sulawesi
The Banggai Islands are an archipelagic district in Central Sulawesi Province, Indonesia, located between mainland Sulawesi and the Maluku Islands. This strategic position places the region within a vital biogeographical transition zone, characterized by a unique mixture of Asian and Australasian flora and fauna. Sulawesi itself is recognized for its exceptional biodiversity and is part of the Wallacea region, a biogeographic zone defined by its distinct ecological characteristics [1]. Similarly, the Maluku Islands exhibit high levels of biodiversity typical of Eastern Indonesia [2]. Consequently, the Banggai Islands serve as an ecological corridor between two biodiversity hotspots, making the area highly significant for conservation and ecological research.
As part of ongoing initiatives to conserve local germplasm and biodiversity, the Kokolomboi Biodiversity Park was established in Banggai Kepulauan Regency [3]. Biodiversity Parks are designated areas outside forest zones that serve both in-situ and ex-situ conservation functions. Initiated by the Indonesian Ministry of Environment and Forestry, this program aims to preserve native or locally adapted plant species facing threats of extinction due to environmental pressures, including land conversion [4]. The park plays a crucial role in maintaining local biodiversity and ensuring the sustainability of key native plant species from the massive land conversion.
Plant species possess distinct adaptive mechanisms that enable them to survive under specific environmental conditions while interacting with other organisms in their surroundings. A critical component of these adaptations involves interactions with root-associated microorganisms, particularly arbuscular mycorrhizal (AM) fungi. These fungi form symbiotic associations with approximately 80–90% of terrestrial plant species [5, 6], contributing significantly to plant health, soil structure, and ecosystem stability [7, 8].
AM fungi enhance plant nutrient uptake, particularly phosphorus, which is often limited in many soils. They also contribute to ecosystem functions by producing glomalin, a glycoprotein that improves soil aggregation and stability [9, 10]. Moreover, AM fungi have demonstrated potential in phytoremediation, especially in soils contaminated with heavy metals [11, 12]. In this mutualistic association, fungal hyphae extend beyond the rhizosphere, accessing water and nutrients unavailable to plant roots, while plants provide carbon compounds necessary for fungal metabolism and growth [10, 13, 14]. This symbiotic interaction contributes to enhancing plant health and its resilience.
The symbiosis between AM fungi and plants plays a key role in vegetation, acting as “a facilitator of communication” within a vegetation community through the formation of a shared mycorrhizal network. This underground network consists of hyphae of AM fungi that physically link the roots of multiple individual plants. Using these networks, individual plants are able to “communicate” and exchange various biochemical signals and resources, including vital nutrients such as phosphorus, secondary metabolites, and defence compounds in response to biotic stressors such as pathogens or herbivores. Additionally, this shared mycorrhizal network enables the transfer of allelopathic substances, which can suppress the growth of competing species, contributing to the underground competition dynamics. The interconnected communication system can enhance adaptability, nutrient efficiency, and ecological interactions among plant communities, thereby influencing the structure and function of the entire ecosystem [15].
The development, colonization, and diversity of AM fungi are influenced by a complex of abiotic and biotic factors, including soil pH, texture, moisture, climate, and the identity of host plant species [10, 14, 16]. Several studies have demonstrated that elevated nitrogen availability in the soil negatively impacts the colonization and growth of AM fungi, as well as the growth of extraradical mycelium. Enhanced soil nutrient availability leads to increased carbon allocation to aboveground plant parts, thereby reducing carbon input to the roots and suppressing AM fungal development [17]. Additionally, the plant’s preference for specific AM fungal taxa can result in dominance of particular fungal species in specific ecosystems [15]. Meanwhile, low AM fungal diversity has been reported in agricultural systems with low plant species diversity [18].
Despite the ecological importance of AM fungi in ecosystem function, research on exploring their diversity and distribution in transitional biodiversity zones such as the Banggai Islands remains scarce. A comprehensive investigation is crucial for understanding the ecological dynamics in complex landscapes. AM fungi are highly sensitive to various soil characteristics and microclimatic conditions, which directly impact their survival, colonization ability, and symbiotic performance. Moreover, the interaction between the AM fungal species and specific vegetation types can determine the diversity of fungi and their function in the ecosystem. It is necessary to closely study the soil characteristics and the interaction between plants and fungi. Therefore, this study aims to assess the diversity and distribution patterns of AM fungal species in relation to vegetation composition and soil properties within the Kokolomboi Biodiversity Park, Banggai Kepulauan Regency, Central Sulawesi, Indonesia. We hypothesized that AM fungal diversity is influenced by vegetation composition and soil nutrients along elevation gradients.
2.1 Research site
The study was conducted in the Kokolomboi Biodiversity Park, located in Leme-Leme Darat Village, Buko District, Banggai Kepulauan Regency, Central Sulawesi, Indonesia (Figure 1). Geographically, this park lies at coordinates 01° 16' 56.40" S and 122° 51' 41.53" E, with a maximum elevation of approximately 1,000 meters above sea level (masl). Sampling plots were established at six locations along an elevational range of 300–800 masl, with three replicate plots at each location. Plots were set at 100-meter elevation intervals to ensure systematic coverage across the gradient.
Figure 1. Research site (red mark) in Kokolomboi Biodiversity Park, Banggai Islands
2.2 Soil and field data collection
Field data collection and soil sampling were conducted using 10 × 10 m plots, following the method of Álvarez-Lopeztello et al. [19]. Within each plot, vegetation data were recorded, and plant specimens were collected for identification at the Herbarium Bogoriense, National Research and Innovation Agency, Indonesia. Soil samples weighing 250 grams were collected from a depth of 20 cm after removing the litter and root layers. The collected soil samples were then transported to the laboratory for the extraction of AM fungal spores and soil chemical analysis.
2.3 Soil analysis
Soil samples were air-dried and gently loosened prior to chemical analysis. Parameters measured included pH measured using potentiometric method, organic carbon (C-organic) measured using the Walkley-Black method, total nitrogen (N) measured using Kjeldahl method, available P₂O₅ measured using Olsen method, potential P₂O₅ and K₂O using HCl 25% extraction method, four nutrients including potassium (K), calcium (Ca), sodium (Na), and magnesium (Mg) measured using atomic absorption spectrophotometer, cation exchange capacity (CEC), and alkaline saturation.
2.4 Spore extraction and identification
AM fungal spores were extracted using the wet sieving and centrifugation method, following the protocol of Walker et al. [20]. A 50-gram subsample of soil was suspended in water and sequentially filtered through 75 μm and 54 μm sieves, followed by centrifugation. The resulting pellets were mixed with a 65% sucrose solution, homogenized using a vortex mixer, and centrifuged again. The supernatant was filtered through a 54 μm sieve, rinsed with water, and the spores were collected in a petri dish for microscopic observation.
Spore mounts were prepared on slides, and for identification, Melzer’s reagent (a mixture of chloral hydrate and potassium iodide) and polyvinyl alcohol–lactic acid–glycerol (PVLG) were used when necessary. Species identification was based on morphological characteristics and followed the keys and references of Schenck [21], Schüßler and Walker [22], the Glomeromycota Phylogeny database (http://www.amf-phylogeny.com), and the International Collection of (Vesicular) Arbuscular Mycorrhizal Fungi (INVAM: https://invam.ku.edu/).
2.5 Statistical analysis
Collected data, including soil variables and vegetation data, were analyzed for correlation. The interpretation of correlation coefficients followed the classification provided by Sugiyono [23]: 0.000–0.199 (very low), 0.200–0.399 (low), 0.400–0.599 (moderate), 0.600–0.799 (strong), and 0.800–1.000 (very strong). The P values were also calculated from the correlation test.
Biodiversity indices, including the Shannon-Wiener diversity index and the Sørensen-Dice similarity index, were calculated using the vegan package (version 2.7-1) [24]. Principal component analysis (PCA) was performed using the Factominer package version 2.11. All graphical visualizations were produced using the ggplot2 package. Statistical analyses and data visualizations were performed using R software version 4.3.1 [25].
3.1 Diversity and distribution of AM fungi
Understanding the diversity and distribution of AM fungi is key to evaluating the ecosystem health and ecological dynamics of plant-microbe interactions, particularly in high-diversity regions. The mutualistic associations between AM fungal species and the roots of terrestrial plants, particularly in nutrient absorption, contribute to the soil structure and stability. In addition, the AM fungi also contribute to supporting the plant community structure, especially in tropical regions where nutrient cycling is dynamic and complex. Documenting the AM fungal diversity offers insight into the potential of soil microbial communities. This is also relevant to the Kokolomboi Biodiversity Park, which has varied elevation, plant communities, and soil nutrients.
The species diversity index calculation indicates that AM fungal diversity across different elevational gradients in this study ranges from a low to moderate range (Table 1). Based on the Shannon-Wiener index (H’), a value less than 1 reflects low diversity. While the value between 1 and 3 indicates moderate diversity [26]. Furthermore, the evenness index values suggest a relatively stable community of AM fungi. The evenness index, which reflects the even distribution of individuals among species, indicates that the value is approaching 1, suggesting a relatively balanced and stable distribution of species within the community, implying ecological stability. The dominance index results indicate a potential dominance by a single species at 801 masl. Notably, only one AM fungus species was recorded at 310 masl, indicating a lack of diversity at this site. In contrast, in mid-elevation sites, species dominance appeared to be low. Species richness, as measured by the Margalef index, presents the highest value at 426 masl, indicating a more favorable condition for AM fungal diversity. These findings suggest that habitat conditions, such as elevation and associated environmental factors, play an important role in shaping the distribution, richness, and abundance of AM fungal species in the study area.
Table 1. Diversity of AM fungi based on elevation
|
Elevation (masl) |
Biodiversity Indices |
Evenness |
Margalef |
Dominance |
||
|
Shannon-Wiener |
Simpson |
InvSimp |
||||
|
310 |
0.0000 |
0.0000 |
1.0000 |
N/A |
0.0000 |
1.0000 |
|
426 |
1.2635 |
0.6578 |
2.9221 |
0.7850 |
0.6201 |
0.3422 |
|
523 |
1.1622 |
0.6446 |
2.8140 |
0.8384 |
0.4651 |
0.3554 |
|
611 |
1.2450 |
0.6797 |
3.1220 |
0.8981 |
0.4651 |
0.3203 |
|
702 |
0.6931 |
0.5000 |
2.0000 |
1.0000 |
0.1550 |
0.5000 |
|
801 |
0.4506 |
0.2778 |
1.3846 |
0.6500 |
0.1550 |
0.7222 |
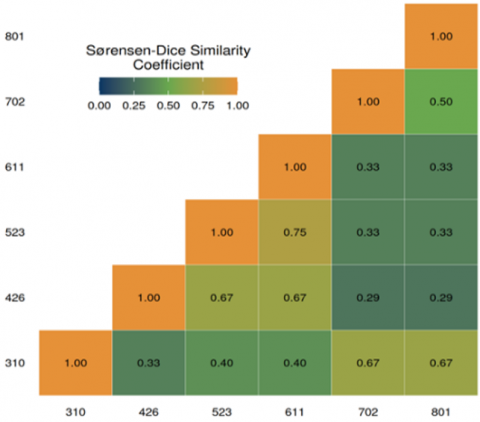
To evaluate the similarity of AM fungal communities across the sites with different elevations, the Sørensen-Dice similarity index was calculated (Figure 2). The results revealed a relatively high degree of community similarity between the sites at 523 and 611 masl. Comparable similarity patterns were also observed among the sites at 426, 523, and 611 masl, as well as between 702 and 801 masl. The Sørensen-Dice index values for the 426–611 masl range (less than 0.5) indicate a low similarity level, whereas the sites at 702–801 masl exhibited index values more than 0.5, suggesting a relatively moderate degree of diversity. These suggest that the minimum elevation differences among these sites create similar microclimatic conditions, such as temperature, humidity, and light intensity, which may contribute to the similar AM fungal community composition. Previous research has shown that environmental factors, including microclimate, water availability, and land-use practices, significantly influence the distribution and abundance of AM fungi [27]. The similarity observed among these elevation ranges may therefore reflect the relative uniformity of these ecological indicators.
Figure 2. The result of the calculation of the Sørensen-Dice similarity coefficient
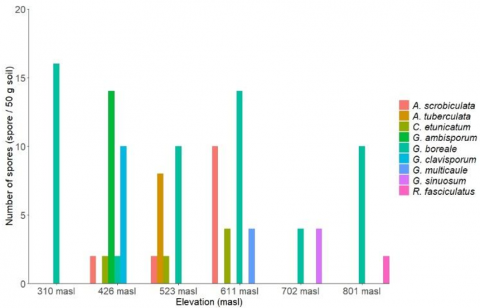
Figure 3. Species of AM fungi at each elevation
A total of nine AM fungal species were identified from six sampling sites within the Kokolomboi Biodiversity Park. These species belonged to four genera: Acaulospora (A. scrobiculata Trappe and A. tuberculata Janos & Trappe), Claroideoglomus [C. etunicatum (W.N. Becker & Gerd.) C. Walker & A. Schüßler], Glomus [G. ambisporum G.S. Sm. & N.C. Schenck, G. boreale (Thaxt.) Trappe & Gerd., G. clavisporum (Trappe) R.T. Almeida & N.C. Schenck, G. multicaule Gerd. & B.K. Bakshi, and G. sinuosum (Gerd. & B.K. Bakshi) R.T. Almeida & N.C. Schenck], and Rhizophagus [R. fasciculatus (Thaxt.) C. Walker & A. Schüßler]. Among them, G. boreale was the only species found across all sampling sites, ranging from 310 to 801 masl (Figure 3). The genus Glomus exhibited the highest species richness in this study, with five species recorded, followed by Acaulospora, which had two species. These findings are consistent with previous studies, such as Datta and Kulkarni [28], who reported that Glomus had the highest relative abundance (75.39%), followed by Acaulospora (8.62%) in sugarcane plantation soils in India. A detailed description of each AM fungal species identified in the Kokolomboi Biodiversity Park is presented below.
Acaulospora scrobiculata Trappe
The spores are rounded, light yellow, and measure approximately 98.97–142.72 μm in width and 99.64–146.97 μm in length . The spore surface is ornamented with fine grooves distributed uniformly across the entire surface. In this study, A. scrobiculata was found at elevations of 426, 523, and 611 masl. It was associated with a diverse range of plant species, including Adinandra celebica, Baccaurea javanica, B. nanihua, Calophyllum sp., Elaeocarpus angustifolius, Ficus septica, Ficus sp., Flacourtia rukam, Glochidion philippicum, Hancea penangensis, Heritiera sylvatica, Kostermanthus heteropetalus, Lithocarpus celebicus, Litsea accedentoides, Pometia pinnata, Premna corymbosa, Prunus arborea, Pterospermum javanicum, Radermachera pinnata, Sophora tomentosa, Syzygium acutangulum, and Vitex cofassus. In other regions of Sulawesi, A. scrobiculata has been reported in association with Acacia mangium, Bothriochloa bladhii, Calopogonium mucunoides, Cyperus sp., Chromolaena odorata, Digitaria longiflora, Desmodium gangeticum, Eupatorium sp., Hyptis capitata, Imperata cylindrica, Leptochloa chinensis, Muntingia calabura, Paspalum conjugatum, Porophyllum ruderale, Uraria lagopodoides, Setaria parviflora [29], and sugarcane Saccharum officinarum [30, 31]. While in Sumba, this species has been associated with bamboo Bambusa spinosa, Chloothamnus reholttumianus, and Schizostachyum brachycladum [32]. In Java, it has been found under Imperata cylindrica (reeds), oil palm (Elaeis guineensis), bamboo, mangosteen (Garcinia mangostana), durian (Durio zibethinus), rambutan (Nephelium lappaceum) [33], and maize (Zea mays) [34].
Acaulospora tuberculata Janos & Trappe
Spores of A. tuberculata are rounded to slightly oval, yellow to orange, and measure approximately 91.9–185.3 μm in width and 94.47–199.63 μm in length. The spore surface is ornamented with small, uniform protrusions that are evenly distributed across the entire surface. In this study, A. tuberculata was found at elevations of 523 and 611 masl. It was associated with a variety of plant species, including Antidesma excavatum, Baccaurea nanihua, Calophyllum sp., Casearia glabra, Citronella suaveolens, Cleistanthus myrianthus, Ficus septica, Flacourtia rukam, Glochidion philippicum, Hancea penangensis, Lithocarpus celebicus, Prunus arborea, Pterospermum celebicum, Radermachera pinnata, Semecarpus heterophyllus, Syzygium acuminatissimum, and S. acutangulum. In Java, A. tuberculata has been reported in association with Theobroma cacao [33, 35]. In Sumba, it has been found colonizing bamboo species such as Dendrocalamus asper, Gigantochloa atter, Schizostachyum brachycladum, and S. lima [32]. In Sulawesi, this species has also been recorded in association with sugarcane (Saccharum officinarum) [30, 31].
Claroideoglomus etunicatum (W.N. Becker & Gerd.) C. Walker & A. Schüßler
Spores of C. etunicatum are rounded, brown, and measure approximately 93.18–122.75 μm in width and 96.76–125.71 μm in length. The subtending hypha is approximately 10.82 μm thick and contains a septum at the spore base, which is characteristic of this species. In this study, C. etunicatum was found at elevations of 426, 523, and 611 masl. It was associated with a wide range of plant species, including Adinandra celebica, Baccaurea javanica, B. nanihua, Calophyllum sp., Elaeocarpus angustifolius, Ficus septica, Ficus sp., Flacourtia rukam, Glochidion philippicum, Hancea penangensis, Heritiera sylvatica, Kostermanthus heteropetalus, Lithocarpus celebicus, Litsea accedentoides, Pometia pinnata, Premna corymbosa, Prunus arborea, Pterospermum javanicum, Radermachera pinnata, Sophora tomentosa, Syzygium acutangulum, and Vitex cofassus.
Glomus ambisporum G.S. Sm. & N.C. Schenck
Spores of G. ambisporum are globose to subglobose and occur in dark brown sporocarps, measuring approximately 119.04–119.40 μm in width and 120.52–126.65 μm in length. In this study, G. ambisporum was recorded at an elevation of 426 masl. It was found in association with several plant species, including Baccaurea nanihua, Ficus sp., Kostermanthus heteropetalus, Premna corymbosa, Pterospermum javanicum, and Vitex cofassus.
Glomus boreale (Thaxt.) Trappe & Gerd.
Spores of G. boreale are oval, brown, and measure approximately 75.14–120.97 μm in width and 101.64–141.22 μm in length. The hyphae connected to the spores are 11.13–18.52 μm thick, while the spore wall thickness ranges from 8.72 to 10.52 μm. This species was recorded across all study sites, at elevations ranging from 310 to 801 masl. Glomus boreale was associated with a broad range of plant species, including Adinandra celebica, Baccaurea nanihua, Calophyllum sp., Canarium littorale, Canarium sp., Chionanthus celebicus, Cleistanthus myrianthus, Elaeocarpus angustifolius, E. culminicola, Endiandra rubescens, Ficus septica, Ficus sp., Glochidion philippicum, Hancea penangensis, Kostermanthus heteropetalus, Lepisanthes fruticosa, Lithocarpus celebicus, Litsea accedentoides, Macaranga hispida, Neonauclea celebica, Palaquium obovatum, Premna corymbosa, Prunus arborea, Pterospermum javanicum, Radermachera pinnata, Santiria laevigata, Saurauia tristyla, Semecarpus heterophyllus, Syzygium acutangulum, S. fastigiatum, Vernonia arborea, and Vitex cofassus.
Glomus clavisporum (Trappe) R.T. Almeida & N.C. Schenck
Sporocarps of G. clavisporum are brown and range in size from 164.00–296.53 μm in width to 171.68–333.89 μm in length. In this study, G. clavisporum was recorded at an elevation of 426 masl. It was associated with several plant species, including Baccaurea nanihua, Ficus sp., Kostermanthus heteropetalus, Premna corymbosa, Pterospermum javanicum, and Vitex cofassus.
Glomus multicaule Gerd. & B.K. Bakshi
Spores of G. multicaule are globose to subglobose, brownish, and measure approximately 207.80 × 225.01 μm. The spores possess a single-layered spore wall and are characterized by the presence of two hyphae attached at opposite sides. This species was recorded at an elevation of 611 masl and was associated with Adinandra celebica, Baccaurea javanica, Heritiera sylvatica, Pometia pinnata, Pterospermum javanicum, and Sophora tomentosa.
Glomus sinuosum (Gerd. & B.K.Bakshi) R.T. Almeida & N.C. Schenck
Sporocarps of G. sinuosum are brown and measure approximately 229.93 × 246.71 μm in diameter. In this study, the species was found at an elevation of 702 masl. It was associated with several plant species, including Ficus septica, Macaranga hispida, Saurauia tristyla, Semecarpus heterophyllus, and Strobocalyx arborea. Additionally, G. sinuosum has previously been reported under oil palm vegetation (Elaeis guineensis) in Java [33].
Rhizophagus fasciculatus (Thaxt.) C. Walker & A. Schüßler
Spores of R. fasciculatus are globose to subglobose, yellow‑brown to brown, and measure approximately 139.20 × 142.96 μm. The spores have three distinct wall layers with a total thickness of 11.62 μm, and the subtending hyphae are approximately 9.61 μm thick. This species was recorded at an elevation of 801 masl and was associated with Baccaurea nanihua, Calophyllum sp., Ficus septica, Glochidion philippicum, Hancea penangensis, Lithocarpus celebicus, Prunus arborea, Radermachera pinnata, and Syzygium acutangulum.
3.2 Correlation between AM fungi and vegetation
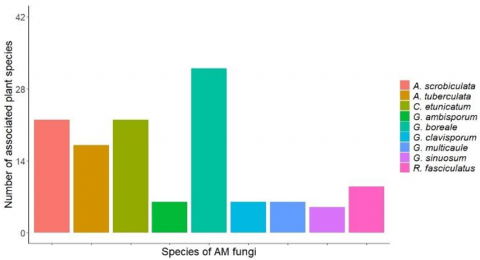
The association between AM fungal species and host plant species varied across taxa (Figure 4). Among all identified AM fungal species, G. boreale exhibited the highest association, being linked with approximately 76% of the 42 plant species observed in the sampling plots. This suggests that G. boreale is capable of forming symbiotic relationships with the majority of tree species present at the study site. Acaulospora scrobiculata and C. etunicatum were each associated with approximately 52.28% of the plant species. Acaulospora tuberculata exhibited a lower association rate, interacting with 40.48% of the plant species, followed by R. fasciculatus, which was associated with 21.43%. Meanwhile, G. ambisporum, G. clavisporum, and G. multicaule each showed similar association frequencies, at 14.29%. The lowest percentage of association was recorded for G. sinuosum, which was linked to only 11.90% of the plant species surveyed.
Figure 4. AM fungal association with plant species in the Kokolomboi Biodiversity Park
AM fungi represent the most widespread type of mycorrhizal symbiosis in nature, forming associations with approximately 80% to 90% of plant species [5, 6]. Accordingly, it is not unexpected that G. boreale, one of the AM fungal species identified in this study, was associated with approximately 76% of the 42 plant species in the observation plots. Previous research has indicated that AM fungal colonization tends to be more prevalent in plant species with poorly developed root systems [36]. Conversely, G. sinuosum exhibited the lowest association rate in this study, interacting with only 11.90% of plant species. This low frequency may be attributed to the influence of environmental factors.
Climatic factors, particularly microclimate conditions, are recognized as a key factor influencing the association between AM fungi and plant species. In this study, the plant species predominantly consist of trees inhabiting forest areas at elevations ranging from 300 to 800 masl. These forests are generally classified as lowland forest ecosystems as defined by Widjaja et al. [37], which typically range in elevations between 0 and 1,000 masl. The lowland forests are characterized by a complex and multi-layer canopy that significantly shapes the microclimatic environment beneath them. The structure creates moderate temperature fluctuation, stable humidity, and decreases light penetration, which supports the growth of diverse soil microbial communities, including AM fungal species. Consequently, the distribution of AM fungi in this ecosystem is likely influenced by soil and microclimate factors, which create the unique microclimatic conditions formed by forest structure and elevation.
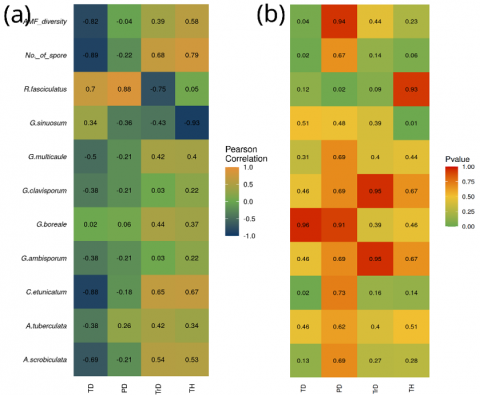
The results of the correlation analysis (Figure 5) indicate the presence of both strong positive and negative correlations between AM fungi and several vegetation types. Specifically, the total number of AM fungal spores and their diversity exhibited strong negative correlations with tree density (TD; r = -0.89 and r = -0.82, respectively) and weak negative correlations with plant species diversity (PD; r = -0.22 and r = -0.04). These findings are consistent with those of Liu et al. [38], who reported a weak relationship between AM fungi and plant species diversity (r = 0.119). However, contrasting evidence has been presented by Chen et al. [39], who found that soils containing a diverse community of AM fungi are more likely to support plant community diversity compared to soils dominated by only one or a few fungal taxa. This suggests that while local correlations may appear weak or negative, the broader ecological role of AM fungal diversity in maintaining plant community structure remains significant.
Moreover, research by Winagraski et al. [16] showed that plant communities often rely on the colonization of AM fungi, with reduced AM fungi abundance linked to the lower plant diversity. This pattern aligns with the findings from the present study, particularly regarding the R. fasciculatus, which was positively correlated with the number of plant species (PD; r = 0.88). Our study revealed that the species of G. sinuosum has a strong correlation with tree height (TH; r = 0,93), suggesting a potential ecological link between this species and plant structure. In contrast, the other four species, namely A. tuberculata, G. ambisporum, G. boreale, and G. clavisporum, exhibited a weak correlation with all measured variables of vegetation characteristics.
3.3 Correlation between AM fungi and soil factors
The establishment and effectiveness of AM fungal symbiosis are also known to be affected by a range of soil and climatic variables, including soil pH, texture, nutrient availability, and the composition of host vegetation [10, 14, 16]. For instance, elevated nitrogen levels have been shown to inhibit the colonization and growth of AM fungal extraradical mycelium [17]. Moreover, increased soil nutrient availability can lead to greater carbon allocation to aboveground plant parts, thereby reducing the carbon supply to the roots and suppressing the development of AM fungi [17].
The results of the chemical analysis of soil samples from the study site (Table 2) were used to further analyze the correlation between soil factors influencing AM fungal communities in the Kokolomboi Biodiversity Park (Figure 6). Most of the sampling sites exhibited slightly acidic to neutral pH (measured in H₂O), and several other soil properties showed minimal variation across locations (Table 2). Soil pH is known to influence the abundance of AM fungal spores, with acidic conditions being considered optimal for spore germination, as AM fungi are generally acidophilic in nature [40].
Table 2. Results of chemical analysis of soil samples from the study site
|
Variable |
Elevation of the Location (meters above sea level) |
|||||
|
310 |
426 |
523 |
611 |
702 |
801 |
|
|
pH H2O |
7.13 (Neutral) |
6.16 (Slightly acidic) |
7.13 (Neutral) |
6.28 (Slightly acidic) |
6.15 (Slightly acidic) |
5.89 (Slightly acidic) |
|
C-organic (%) |
19.27(5) |
16.54(5) |
16.02(5) |
6.14(5) |
17.01(5) |
16.64(5) |
|
N-total (%) |
1.01(5) |
0.75(4) |
0.75(4) |
0.42(3) |
0.60(4) |
0.57(4) |
|
C/N ratio |
19.08(4) |
22.05(4) |
21.36(4) |
14.61(3) |
28.35(5) |
29.19(5) |
|
Available P2O5 (mg/kg) |
23.81(5) |
19.45(4) |
44.19(5) |
57.94(5) |
46.20(5) |
29.62(5) |
|
Potential P2O5 (mg/100 g) |
23.80(3) |
13.00(1) |
22.77(3) |
54.63(4) |
27.19(3) |
18.43(2) |
|
Potential K2O (mg/100 g) |
14.44(2) |
17.11(2) |
13.65(2) |
7.76(1) |
8.23(1) |
11.42(2) |
|
K (cmol/kg) |
0.20(2) |
0.25(2) |
0.14(2) |
0.10(2) |
0.10(2) |
0.19(2) |
|
Na (cmol/kg) |
0.07(1) |
0.04(1) |
0.10(2) |
0.05(1) |
0.01(1) |
0.26(2) |
|
Ca (cmol/kg) |
50.26(5) |
30.11(5) |
56.30(5) |
8.54(3) |
36.21(5) |
4.87(2) |
|
Mg (cmol/kg) |
1.72(3) |
2.41(4) |
2.16(4) |
0.84(2) |
1.31(3) |
0.79(2) |
|
Cation exchange capacity (cmol/kg) |
43.24(5) |
39.51(4) |
37.66(4) |
18.57(3) |
28.08(4) |
22.87(3) |
|
Alkaline saturation (%) |
100(5) |
83.02(5) |
(100(5) |
51.21(3) |
100(5) |
26.69(2) |
The abundance and diversity of AM fungi do not always have a very strong correlation with soil nutrient levels. This finding contrasts with other research results, which have shown that the availability of soil nutrients for plants can stimulate AM fungal sporulation and increase spore populations. Conversely, spore populations decrease when the nutrient availability is decreased [42]. For instance, nutrients N and P in an ecosystem affect the abundance of AM fungi, especially their diversity [43].
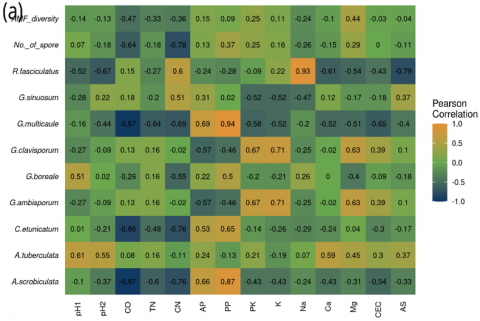
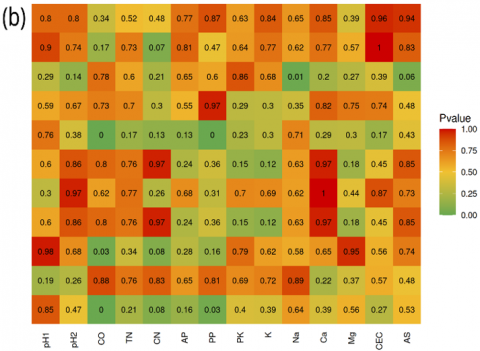
The species A. scrobiculata and G. multicaule have positive correlations with P2O5 potential (PP; r = 0.87 and r = 0.94) and negative correlations with C-organic (CO; r = -0.97). The C. etunicatum was negatively correlated with tree density (TD; r = -0.88) and C-organic (CO; r = -0.86). These findings align with Soka and Ritchie’s results, which indicate that variations in the distribution of AM fungi are influenced by habitat preferences and differing levels of tolerance to elevated soil nutrient concentrations [44]. As a result, the abundance of AM fungal species may decline when soil nutrient levels exceed optimal levels.
It is well established that the symbiosis, development, and diversity of AM fungi in an area are impacted by soil factors (soil pH and soil type), host plant species, and climate conditions [10, 14, 16]. The weak correlations observed for these four species suggest that their abundance and distribution are likely to be more closely tied to specific microclimate condition factors or unmeasured ecological factors.
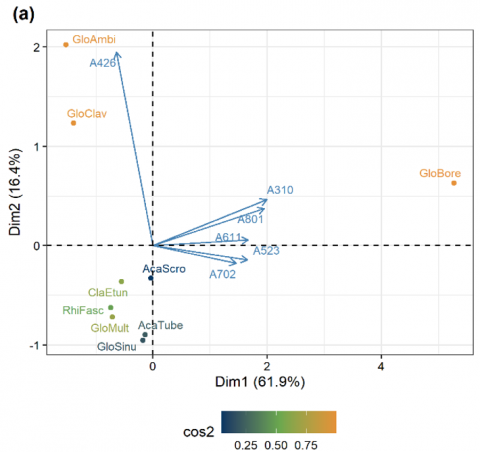
To understand the influence of environmental factors, particularly plot height, on the abundance of AM fungal species, a PCA was conducted. The results of this analysis are presented in Figure 7.
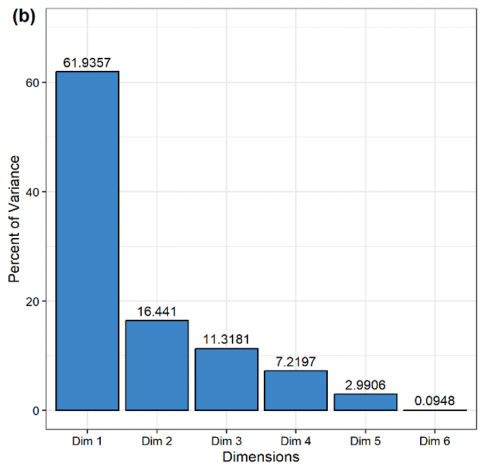
The squared cosine (cos2) value from the PCA shows that Glomus boreale (cos2 0.98 at Dim1) is the most important AM fungus in the observed community. Furthermore, A. tuberculata and G. sinuosum are AM fungal species with low cos2 values because they are only found in 1 plot with low abundance. The high cos2 value indicates that PCA well represents these AM fungal species [45, 46]. The eigenvalue shows that Dim1 in PCA can explain more than 50% of the variance in the analyzed data (Figure 7). Dim1 represents the elevation gradients except the elevation of 426 masl, which is well represented by Dim2. Based on Figure 7, a significant correlation is also seen between the plot at an elevation of 310 masl (A310) and the plot at an elevation of 801 masl (A801). Likewise, the plots at elevations of 523 and 702 masl. The abundance of AM fungi in each plot also influences the correlation between the plots. The directions and length of the arrows indicate the correlation and contribution of each variable to Dim1 and Dim2. In this study, we found that elevation of 310 masl is the most contributing variable for Dim1, followed by elevation of 801 masl and elevation of 611 masl, respectively.
This study identified nine species of AM fungi that are associated with various plant communities in the Kokolomboi Biodiversity Park. These species included Acaulospora scrobiculata, A. tuberculata, Claroideoglomus etunicatum, Glomus ambisporum, G. boreale, G. clavisporum, G. multicaule, G. sinuosum, and Rhizophagus fasciculatus. The G. boreale emerged as the most widely distributed species, occurring across all sampling sites and exhibiting associations with the majority of plant species, highlighting its ecological adaptability. In contrast, G. sinuosum was the least frequently encountered. Furthermore, five AM fungal species demonstrated a very strong correlation with one or more vegetation characteristics and/or soil factors. These findings emphasize that both plant composition and soil conditions significantly influence the abundance and distribution of AM fungal communities in the study area. This study provides information on AM fungi that contribute to strengthening ecosystem functions.
The authors would like to thank Dr. Kartini Kramadibrata, who provided revisions and suggestions in the preparation of the manuscript, Idang Sumanta, Supardi Jakalalana, M. Syarifudin Hidayatulah, Deni Sahroni, and those who have helped collect data in the field, identify plant species, and analyze them in the laboratory.
[1] Bashari, H., Cryan, M., Dwisasanti, N., Hermansyah, A., Pardede, S., Saryanthi, R., Sasmitawidjaja, V., Savitri, L., Susandi, A., Susanti, T., Tetuka, B., Walshm, T., Wood, P. (2014). Hotspot Keragaman Hayati Wallacea. Critical Ecosystem Partnership Fund, 3-39. https://www.cepf.net/resources/ecosystem-profile-documents/wallacea-ecosystem-profile-summary-0.
[2] Rumanta, M., Lelloltery, H., Kunda, R.M., Kakisina, P. (2019). Selection of plant species as feed sources and nesting places salmon-crested cockatoo (Cacatua moluccensis) Maluku endemic in Manusela National Park (MNP). Advances in Animal and Veterinary Sciences, 7(6): 474-479. https://doi.org/10.17582/journal.aavs/2019/7.6.474.479
[3] Setiadi, A., Khasanah, S.N., Alhumaira, F., Zafirah, T. (2024). An integration of community empowerment and biodiversity conservation program through social-ecology approach in Indonesia (study case: Kokolomboi Hamlet). Journal of Sustainable Development, 17(2): 84-95. https://doi.org/10.5539/jsd.v17n2p84
[4] Indonesian Government. (2012). Regulation of the Minister of State for Environment of the Republic of Indonesia Number 03 of 2012. Minister of Law and Human Rights of the Republic of Indonesia. https://jdih.menlhk.go.id/kiosk/files/MLH%20P.3%20(1).pdf.
[5] Hartini, H. (2020). Exploration of the potential of Siam weed (Chromolaena odorata) as compost and arbuscular mycorrhzal fungi (AMF) on the growth of cacao seedlings. Journal of Media Agrosains, 6(1): 7-13. https://repository.polteklpp.ac.id/id/eprint/3410/.
[6] Kramadibrata, K. (2013). Diversity of Glomeromycota in Baturaden Botanic Garden and its surrounding area in Slamet Mountain. Berita Biologi, 12(2): 217-222.
[7] Bhupenchandra, I., Chongtham, S.K., Devi, A.G., Dutta, P., et al. (2024). Unlocking the potential of arbuscular mycorrhizal fungi: Exploring role in plant growth promotion, nutrient uptake mechanisms, biotic stress alleviation, and sustaining agricultural production systems. Journal of Plant Growth Regulation. https://doi.org/10.1007/s00344-024-11467-9
[8] Chaudhary, A., Poudyal, S., Kaundal, A. (2025). Role of arbuscular mycorrhizal fungi in maintaining sustainable agroecosystems. Applied Microbiology, 5(1): 6. https://doi.org/10.3390/applmicrobiol5010006
[9] Koziol, L., Schultz, A.P., House, G.L., Baurer, J.T., Middleton, E.L., Bever, J.D. (2018). The plant microbiome and native plant restoration: The example of native mycorrhizal fungi. BioScience, 68(12): 996-1006. https://doi.org/10.1093/biosci/biy125
[10] Ullah, S., Muhammad, B., Amin, R., Abbas, H., Muneer, M.A. (2019). Sensitivity of arbuscular mycorrhizal fungi in old-growth forest: Direct effect on growth and soil carbon storage. Applied Ecology and Environmental Research, 17(6): 13749-13758. https://doi.org/10.15666/aeer/1706_1374913758
[11] Zulkoni, A., Rahyuni, D., Nasirudin, N. (2017). Under ground root pruning and AMF inoculation to improve phytoremediation of soil contaminated with mercury due to gold mining by Tectona grandis in Kokap Kulonprogo Yogyakarta. Jurnal Manusia dan Lingkungan, 24(1): 17-22. https://doi.org/10.22146/jml.23071
[12] Zulkoni, A. (2018). Efforts to improve phytoremediation of mercury-contaminated soil in Kokap Kulonprogo Yogyakarta using Acacia (Acacia Sieberiana DC) with root pruning and Mycorrhza inoculation. Jurnal Rekayasa Lingkungan, 18(1): 1-10. https://doi.org/10.37412/jrl.v18i1.19
[13] Suryati, T. (2017). Study of Arbuscular Mycorrhizal fungi in tin post-mining land of Central Bangka Regency. Jurnal Teknologi Lingkungan, 18(1): 45-53.
[14] Wang, J., Wang, G.G., Zhang, B., Yuan, Z., Fu, Z., Yuang, Y., Zhu, L., Ma, S., Zhang, J. (2019). Arbuscular mycorrhizal fungi associated with tree species in a planted forest of Eastern China. Forests, 10(5): 424. https://doi.org/10.3390/f10050424
[15] Gorzelak, M.A., Asay, A.K., Pickles, B.J., Simard, S.W. (2015). Inter-plant community through mycorrhizal networks mediates complex adaptive behaviour in plant communities. AoB Plants, 7: pvls050. https://doi.org/10.1093/aobpla/plv050
[16] Winagraski, E., Kachuk, G., Monteiro, P.H.R., Auer, G.C., Higa, A.R. (2019). Diversity of arbuscular mycorrhizal fungi in forest ecosystems of Brazil: A review. Cerne, 25(1): 25-35. https://doi.org/10.1590/01047760201925012592
[17] Milovic, M., Kebert, M., Orlovic, S. (2021). How mycorrhizas can help forests to cope with ongoing climate change? Šumarski List, 145(5-6): 279-286. https://doi.org/10.31298/sl.145.5-6.7
[18] Corryanti, C. (2011). Arbuscular mycorrhizal fungi in teak plantations intercropped with sugarcane. Jurnal Agrotropika, 16(1): 1-8. https://doi.org/10.23960/ja.v16i1.4259
[19] Álvarez-Lopeztello, J., Castillo, R.F.D., Robles, C., Hernandez-Cuevas, L.V. (2019). Spore diversity of arbuscular mycorrhizal fungi in human-modified neotropical ecosystem. Ecological Research, 34(3): 394-405. https://doi.org/10.1111/1440-1703.12004
[20] Walker, C., Mize, C.W., McNabb Jr., H.S. (1982). Populations of endogonaceous fungi at two locations in central Iowa. Canadian Journal of Botany, 60(12): 2518-2529. https://doi.org/10.1139/b82-305
[21] Schenck, N.C. (1990). Manual for the Identification of VA Mycorrhizal Fungi, 3rd ed. Synergistic Publications Gainesville.
[22] Schüßler, A., Walker, C. (2010). The Glomeromycota. A species list with new families and new genera. Gloucester, UK. https://www.researchgate.net/publication/285775809_The_Glomeromycota.
[23] Sugiyono, P.D. (2012). Metode Penelitian Kuantitatif Kualitatif dan R&D, Cet. Ke-12 [Qualitative and Quantitative Research Methods and R&D, 12th edition]. Bandung: Alfabeta.
[24] Oksanen, J., Simpson, G.L., Blanchet, F.G., Kindt, R., et al. (2022). Vegan: Community Ecology Package: Ordination methods, diversity analysis and other functions for community and vegetation ecologists. R Package Version 2.7-1. https://doi.org/10.32614/CRAN.package.vegan
[25] TRF (The R Foundation). (2021). The R Project for Statistical Computing. https://www.r-project.org.
[26] Adelina, M., Harianto, S.P., Nurcahyani, N. (2016). Bird diversity in the community forest of Desa Kelungu, Sub-district Kotaagung, Tanggamus District. Jurnal Sylva Lestari, 4(2): 51-60. https://doi.org/10.23960/jsl2451-60
[27] Jamiołkowska, A., Księżniak, A., Gałązka, A., Hetman, B., Kopacki, M., Skwaryło-Bednarz, B. (2018). Impact of abiotic factors on development of the community of arbuscular mycorrhizal fungi in the soil: A review. International Agrophysics, 32(1): 133-140. https://doi.org/10.1515/intag-2016-0090
[28] Datta, P., Kulkarni, M. (2012). Arbuscular mycorrhizal fungal diversity in sugarcane rhizosphere in relation with soil properties. Notulae Scientia Biologicae, 4(1): 66-74. https://doi.org/10.15835/nsb416567
[29] Tuheteru, F.D., Husna, Albasri, Arif, A., Wulan, S.A., Kramadibrata, K. (2019). Arbuscular mycorrhizal fungi associated with adaptive plants in gold mine tailing. Biodiversitas, 20(11): 3398-3404. https://doi.org/10.13057/biodiv/d201137
[30] Kumalawati, Z., Musa, Y., Amin, N., Asrul, L., Ridwan, I. (2014). Exploration of arbuscular mycorrhizal fungi from sugarcane rhizosphere in South Sulawesi. International Journal of Scientific & Technology Research, 3(1): 201-203.
[31] Kumalawati, Z. (2015). Identifikasi dan isolasi spora tunggal cendawan mikoriza arbuskula pada rhizospheren tebu (Saccharum officinarum L.) [Identification and isolation of single spores of arbuscular mycorrhizal fungi in the rhizosphere of sugarcane (Saccharum officinarum L.).]. In Prosiding Seminar Nasional Biologi, pp. 63-71.
[32] Kramadibrata, K. (2011). Diversity of bamboo JA on Sumba Island. Berita Biologi, 10(5): 635-639.
[33] Kramadibrata, K. (2009). Glomeromycota recovered from cacao soil. Reinwardtia, 12(5): 357-371.
[34] Haerida, I., Kramadibrata, K. (2002). Identification of arbuscular mycorrhizal fungi in the rhizosphere of sweet corn plants in Java. Floribunda, 2(2): 33-37. https://www.ptti.or.id/journal/index.php/Floribunda/article/view/38.
[35] Kramadibrata, K. (2016). Diversity of Arbuscular Fungi in Enggano Island. Berita Biologi, 15(3): 59910.
[36] Kramadibrata, K., Gunawan, A.W. (2006). Arbuscular mycorrhizal fungi surrounding tropical kudzu and para grass. Jurnal Mikrobiologi Indonesia, 11: 97-102.
[37] Widjaja, E.A., Rahayuningsih, Y., Rahajoe, J.S., Ubaidilla, R., Maryanto, I., Walujo, E.B., Semiadi, G. (2014). Kekinian Keanekaragaman Hayati Indonesia 2014. LIPI Press. https://penerbit.brin.go.id/press/catalog/view/20/19/22126.
[38] Liu, L., Zhu, K., Wurzburger, N., Zhang, J. (2020). Relationships between plant diversity and soil microbial diversity vary across taxonomic groups and spatial scales. Ecosphere, 11(1): e02999. https://doi.org/10.1002/ecs2.2999
[39] Chen, M., Arato, M., Borghi, L., Nouri, E., Reinhardt, D. (2018). Beneficial services of arbuscular mycorrhizal fungi-from ecology to application. Frontiers in Plant Science, 9: 1270. https://doi.org/10.3389/fpls.2018.01270
[40] Sianturi, F., Linda, R., Khotimah, S. (2005). Density of arbuscular vesicular mycorrhizal fungi spores at three levels of peat maturity in the Mount Ambawang Protected Forest Area, Kubu Raya Regency. Protobiont, 4(2): 96-102. https://jurnal.untan.ac.id/index.php/jprb/article/view/11785.
[41] Eviati, Sulaeman, Herawaty, L., Anggria, L., Usman, Tantika, H.E., Prihatini, R., Wuningrum, P. (2023). Technical guidelines for chemical analysis of soil, plants, water and fertilizers. Kementerian Pertanian Republik Indonesia. https://repository.pertanian.go.id/handle/123456789/24650.
[42] Nandi, R., Mridha, M.A.U., Bhuiyan, M.K. (2014). Seasonal dynamics of arbuscular mycorrhizal fungi (AMF) in forest tree of Chittagong University campus in Bangladesh. Journal of Forest and Environmental Science, 30(3): 277-284. https://doi.org/10.7747/JFS.2014.30.3.277
[43] Camenzind, T., Hempel, S., Homeier, J., Horn, S., Velescu, A., Wilcke, W., Rilig, M.C. (2014). Nitrogen and phosphorus additions impact arbuscular mycorrhizal abundance and molecular biodiversity in a tropical montane forest. Global Change Biology, 20(12): 3646-3659. https://doi.org/10.1111/gcb.12618
[44] Soka, G., Ritchie, M. (2014). Arbuscular mycorrhizal symbiosis and ecosystem processes: Prospects for future research in tropical soils. Open Journal of Ecology, 4(1): 11-22. http://doi.org/10.4236/oje.2014.41002
[45] Priyadharshini, Radha, M., Kumaraperumal, R., Vanitha, G., Kannan, B. (2021). Characterization of environmental covariates of Coimbatore district using principal component analysis. International Journal of Current Microbiology and Applied Sciences, 10(1): 3114-3123. https://doi.org/10.20546/ijcmas.2021.1001.362
[46] Bayraklı, B., Dengiz, O., Erkoçak, A. (2025). Assessment of enzyme activities related to heavy metal pollution risks in long-term hazelnut cultivation soils under humid ecosystem conditions. Geomicrobiology Journal, 42(6): 459-475. https://doi.org/10.1080/01490451.2025.2471469