GC-MS Characterization of Antibacterial Compounds from Bacillus thermoamylovorans Isolated from Human Skin Microbiome

Manal K. Mohammad*![]() | Suzan S. Hussain
| Suzan S. Hussain![]() | Amer S. Mahdi
| Amer S. Mahdi![]()
© 2025 The authors. This article is published by IIETA and is licensed under the CC BY 4.0 license (http://creativecommons.org/licenses/by/4.0/).
OPEN ACCESS
In this study, bacteria from the genus Bacillus were selected to produce new antibiotics required at the therapeutic level due to the high resistance shown by pathogens to current antibiotics. We worked on 50 isolates of this type of bacteria taken from the skin of healthy individuals. We were able to identify 9 isolates from them. Antibacterial activity of the nine isolates was evaluated against bacteria causing urinary tract diseases, where Bacillus thermoamylovorans showed the strongest antimicrobial activity and was identified as a new spore-forming bacterium against most test organisms compared to other Bacillus isolates, followed by B. subtilis and B. amyloliquefaciens, particularly against P. aeruginosa and E. coli. The purified fractions F13-F16 demonstrated inhibition rates ranging from 23.33% to 89.67% depending on the strain of bacteria, with F15 showing the highest inhibition rate up to 86.00% against Klebsiella pneumoniae. The optimum growth conditions were determined at 50℃ and pH 7, and continued for 96 hours in tryptic soy broth for Bacillus thermoamylovorans. The ideal carbon source for growth was determined to be 4% glucose. To extract the antimicrobial metabolites, ethyl acetate solution (1:1 v/v) was used. The concentrated bacterial extract was obtained, and after confirming its biological effectiveness against bacteria causing urinary tract diseases, it was purified using column chromatography to separate the active components into a number of tubes. Further investigation was conducted on the antimicrobial activity of the collected tubes against a group of Gram-negative and Gram-positive pathogenic bacteria, as well as uropathogenic bacteria. The effective tubes were identified from the others. Samples of the active tube solutions were sent for analysis using GC-MS to identify the active compounds through their high relative concentrations, such as trans monounsaturated fatty acids (cis-vaccenic acid), monounsaturated fatty acids (oleic acid - 9-octadecenoic acid), saturated fatty acids (pentadecanoic acid, myristic acid), and polyunsaturated fatty acids (linoleic acid). These fatty acids have known anti-inflammatory, antibacterial, and antioxidant activities, that to the overall bioactivity observed. The most important objectives of this study are to develop new alternatives in antimicrobial strategies.
Bacillus thermoamylovorans, ethyl acetate, uropathogenic bacteria, GC-MS
The rise of antibiotic resistance among pathogenic bacteria necessitates the exploration of alternative therapeutic strategies [1]. One promising avenue is the utilization of substances derived from the human microbiota to inhibit the overgrowth of pathogenic competitors. Despite this potential, there has been limited analytical research focusing on the competitive dynamics among human microorganisms for drug discovery and development [2].
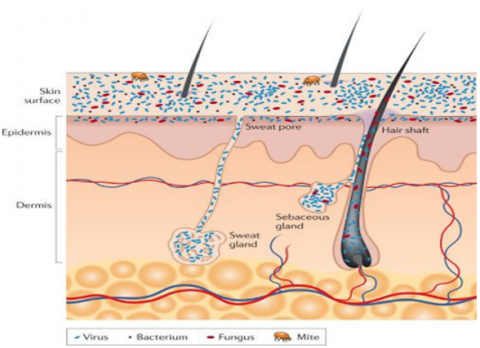
The skin in the human body is the largest organ, harboring diverse bacterial communities that interact with both their host and one another. These skin-resident bacteria play a crucial role in protecting against infections by pathogenic microorganisms due to long-term co-evolution between the skin-resident microorganisms and the human host. It has promoted the specific antagonistic compounds production that is useful to maintain microbial balance and prevent the colonization of pathogens, highlighting the skin as a promising source of novel antibiotics. Microorganisms (bacteria, viruses, fungi, and mites) cover the skin surface, and in the hair and glands reside deep. On the surface of the skin, round and rod-shaped bacteria such as Staphylococcus spp. and Proteobacteria, respectively, form communities that are intertwined among themselves and deeply embedded among other microorganisms. As individual cells and as branching filamentous hyphae, commensal fungi such as Malassezia spp. grow both. Particles of the virus live both in bacterial cells and freely. Skin mites, like Demodex brevis and Demodex folliculorum, are some of the smallest arthropods and live near or in hair follicles. Appendages of skin include sebaceous glands, hair follicles, and sweat glands (Figure 1) [3].
Figure 1. Skin histology schematic in cross-section, viewed with skin appendages and microorganisms
For instance, certain strains can thrive at elevated temperatures (between 40℃ and 58℃), making those candidates for novel antimicrobial agents.
Notably, Bacillus thermoamylovorans has been identified as a source of antimicrobial structures that could combat pathogenic bacteria [4]. Bacillus thermoamylovorans is a thermophilic bacterium that can be isolated from various environments, including skin. Makes it of interest in both industrial applications and microbiological research, with its ability to survive high temperatures and produce enzymes such as lipases.
To their production of various compounds, including antibiotics and enzymes, the widespread presence and metabolic versatility of Bacillus species contribute significantly. This makes them particularly interesting for applications in biotechnology, such as engineered industrial enzymes used in food, detergent industries, and the production of fermented foods. Bacillus species are known sources of secondary metabolites as bioactive compounds exhibiting antifungal, antibacterial, and anticancer properties. Secondary metabolites (SM) are produced as organic compounds from different organisms, especially plants, fungi, and bacteria.
Primary metabolites, which are involved directly in growth and reproduction, while secondary metabolites play essential roles in ecological interactions, defense mechanisms, and have potential therapeutic applications. Their mechanisms of action are diverse and can involve multiple pathways and targets [5]. Given their safety profile and eco-friendly nature, secondary metabolites from bacteria are increasingly recognized as viable alternatives to synthetic drugs. This understanding can guide further modifications to optimize efficacy. In conclusion, as antibiotic resistance continues to pose significant challenges in healthcare, leveraging the unique properties of skin microbiota and employing advanced techniques like molecular docking could pave the way for developing novel antimicrobial agents and secondary metabolites derived from bacteria, offering promising opportunities for treating microbial infections, with multiple benefits including reduced resistance and enhanced therapeutic efficacy [6].
This study aims to isolate and identify bioactive antibiotic compounds from Bacillus thermoamylovorans, a thermophilic bacterium, and evaluate their antibacterial efficacy against skin microbiome-associated pathogens. Gas Chromatography-Mass Spectrometry (GC-MS) will be used to analyze and characterize the chemical composition of these compounds, exploring their potential as novel antibiotic agents for treating skin infections or modulating the skin microbiome.
2.1 Isolation of Bacillus spp.
Skin swab samples were collected from areas that may harbor thermophilic bacteria, such as the forearm, foot, and neck. Ensure that the area is not treated with antimicrobial agents prior to sampling to avoid inhibiting bacterial growth. Fifty samples were collected from the skin of healthy humans aged 13 to 50 years, and the skin swabs were suspended in 100 mL of sterile saline. This suspension should be gently agitated to dislodge bacteria from the swab. Spread the suspension onto selective media such as tryptic soy agar (TSA) plates.
Incubate the plates at elevated temperatures of 50℃ for 24 h to promote the growth of thermophilic organisms while suppressing mesophilic bacteria. After incubation, examine the plates for colonies that exhibit the characteristic morphology of Bacillus species (e.g., creamy white circular colonies). Select colonies that appear homogeneous and purify them by streaking on fresh agar plates. Bacillus thermoamylovorans can be characterized based on microscopic examination, morphological features, and biochemical tests using Bergey’s Manual as a reference. Identification was further supported by the VITEK 2 analyzer with a colorimetric reagent card that was used (BioMérieux, France) for species-level assignment, with identification confidence thresholds set at > 90%.
Additionally, 16S rRNA sequencing was performed and similar to sequences of reference organisms using the NCBI Basic Local Alignment Search Tool (BLASTn). The strains were purified and stored on nutrient agar slants at 4℃ for two months at -20℃ in glycerol for longer-term storage 6.
Performed PCR to amplify the 16S rRNA gene using the following components in Table 1.
Table 1. PCR reaction components for 16S rRNA gene amplification
|
Component |
Volume |
|
Trans gen Biotech (Master mix Easy Taq (TRN)) |
12.5 µl |
|
Forward primer |
0.5 µl |
|
Reverse primer |
0.5 µl |
|
DNA template |
5 µl |
|
Free nuclease water |
6.5 µl |
2.2 Genomic DNA isolation by the boiling method
To select the strain, DNA was isolated using the boiling method. The isolate was streaked onto a plate of fresh agar, and approximately 10 colonies were transferred into an Eppendorf tube containing 400 μl of double-distilled water (dd H₂O). To lyse the cells, take the Eppendorf tube and place it in a water bath at 100℃ for 15 min. The tube was then cooled directly on ice and subjected to heat shock at -20℃ for 20 min. Afterward, the tube was allowed to stand at room temperature and homogenized by vortexing for 10 s. The mixture was centrifuged for 15 min at 4℃. The upper layer was aqueous, which was carefully collected and taken into another sterile Eppendorf tube, and this DNA sample was frozen until further use [7].
PCR conditions used as follows:
1.94℃ for 5 min as the initial denaturation of DNA
2.94℃ for 30 s as the denaturation step of DNA
3.55℃ for 30 seconds as the annealing step of the primers
4.72℃ for 1 min 30 seconds - as the elongation step. 35 cycles of step 2-4 were performed.
5.72℃ for 10 min as the final extension step
2.2.1 DNA agarose gel electrophoresis
The products of PCR were examined using agarose gel electrophoresis. A prepared 1% agarose gel was made by melting 0.5 g of agarose in 50 ml of 1x TBE (Tris-Borate-EDTA) buffer. Before adding ethidium bromide, the agarose was allowed to cool to a temperature of 50-60℃ to reach a concentration of 0.5 μg/ml. After the gel was poured into a gel tray and a comb, the gel was left to solidify at room temperature.
Load the samples into the gel alongside a DNA marker to determine accurately the size of the fragments of the DNA. The gels were run in 1x TBE buffer at a constant voltage of 100 V. DNA bands were visualized using an ultraviolet transilluminator imaging system.
Primers used:
-Forward Primer: Eu8f – AGAGTTTGATCCTGGCTCAG
-Reverse Primer: Eu1492r - GGCTACCTTGTTACGACTT
2.3 Preparation of tester strains
One hundred bacterial isolates were obtained from clinical specimens, specifically from urinary tract infections, which were obtained from different hospitals in Baghdad. The isolates were cultured on suitable media for re-identification. The identified isolates included Escherichia coli, Staphylococcus aureus, Klebsiella pneumoniae, and Pseudomonas aeruginosa. Identification was based on morphological and cultural characteristics of the colonies, which were assessed through bacterial growth on agar culture media, microscopic examination, biochemical tests, and the compact system of VITEK 2.
The automated system VITEK 2 is a bacterial identification and susceptibility testing system that provides rapid results. It employs fluorescence-based technology to accurately identify bacteria and assess their antibiotic susceptibility. For both Gram-negative and Gram-positive bacteria studies have shown that the VITEK 2 system can achieve high rates of correct identification, for susceptibility testing reaching up to 96 with overall agreement rates [8], this study exhibited high resistance of isolates from both types of bacteria to antibiotics were selected to test the effect of bacterial extracts on them, this selection process is for understanding crucial the efficacy of potential antimicrobial agents against resistant strains.
2.4 Antimicrobial activity determination of Bacillus spp. using spectrophotometer
Bacillus spp., which recorded a producer isolate to antimicrobial activity against uropathogenic bacterial strains, and evaluated the extract by the spectrophotometric method [9]. By measuring optical density (OD) at specific wavelengths, this approach allows for precise quantification of bacterial growth inhibition. For preparation of bacterial extracts from Bacillus spp. and preparation of pathogenic bacteria as tested strains for evaluate the extract against it tryptic soy broth (TSB) inoculated by suspension containing (9 ×108 CFU /ml) (compared to McFarland standard 0.5 ml absorbance at 600 nm a wavelength about 0.134) with size 2% inoculum and incubated at 37℃ for 24 h until reaching the stationary phase. After incubation, the culture medium was centrifuged at 10000 rpm for 10 min, the supernatants were collected to obtain the bacterial extracts, and removed the biomass then the supernatant used to estimate activity of the bioactive compounds by added crude extract 0.5 ml to test tubes containing 0.5 ml standardized inoculated of organisms, involving Escherichia coli, Staphylococcus aureus, Klebsiella pneumoniae and Pseudomonas aeruginosa. At 37℃ for 24 h, each tube was incubated, and the optical density (OD) of each culture was measured using a spectrophotometer at 600 nm (OD 600). The reduction in OD compared to control cultures (without extract) indicated the level of antimicrobial activity [10, 11].
Inhibition Rate Calculation Equation:
To calculate the inhibition rate of bacterial extracts, the following equation can be used:
Inhibition Rate = ((ODcontrol - ODsample) / ODcontrol) × 100% (1)
where, ODcontrol = Optical density of the control sample (without extract) was prepared by adding 0.5 ml D.W. and 0.5 ml standardized inoculum of organisms. ODsample = Optical density of the sample containing the extract. was prepared by adding 0.5 ml crude extract to 0.5 ml standardized inoculum of organisms. The blank was prepared by adding 0.5 ml D.W. to 0.5 ml TSB.
2.5 Effect of growth media, pH, temperature, incubation period, and glucose concentration on the production of active compounds from Bacillus thermoamylovorans
To investigate the optimal conditions for the maximum production of antibacterial compounds from Bacillus thermoamylovorans, various factors were examined, including growth media types, incubation temperature, incubation periods, pH levels, and glucose concentrations. This study followed the methodology outlined in the work [11].
Experimental Conditions
Temperature: The strain was cultured at temperatures of 25℃, 30℃, 37℃, and 50℃ for varying periods of 24, 48, 72, 96, and 144 h. Bacillus thermoamylovorans is known to grow optimally between 40℃ and 58℃ and thrives at higher temperatures.
pH Levels: Based on the work [12], the pH was adjusted to values of 5, 6, 7, 8, and 9. Generally favorable for the growth of many Bacillus species is the neutral pH of around 7.
Growth Media: For culturing the bacterium, different media were utilized, including tryptic soy and nutrient broth, and brain heart infusion. Each medium provides distinct nutrients that can affect microbial growth and metabolite production [13].
Glucose Concentration: Various concentrations of glucose (1%, 2%, 3%, 4%, and 5%) were added to the optimal growth medium. Glucose serves as a primary carbon source that can significantly influence the metabolic activity of Bacillus spp., promoting the production of antimicrobial compounds [14]. After optimizing the culture conditions at pH 7 and a temperature of 50℃ for four days, we centrifuged each culture at 3000 rpm for 20 min to separate the supernatant. Evaluated antimicrobial activity of the isolates against Staphylococcus aureus by a broth macrodilution assay as described in CLSI standard guidelines.
2.6 Cultivation and extraction of antimicrobial compounds
Bacillus thermoamylovorans was cultivated in 500 ml of tryptic soy broth, inoculated with 5% active culture, and maintained at 200 rpm at 50℃ for 4 days. The process of fermentation was conducted according to the optimal conditions established by the study [12].
After the incubation period, filtered the fermentation broth through filter paper, Whatman No. 1, to remove cellular debris, remove then centrifuged the filtrate at 10.000 rpm at 4℃ for 20 min to remove the supernatant from the pellet.
For the extraction of antibacterial compounds, the culture filtrate was mixed twice with an equal volume of ethyl acetate (1:1). The phase of ethyl acetate that had the active compounds from the aqueous phase was separated using a separating funnel. Concentrated ethyl acetate layer by evaporating it at 50℃, resulting in a brown crude extract weighing approximately 1.8 g [14]. For antibacterial activity against uropathogenic bacteria, the extracted compounds were bioassayed using standard methods.
2.6.1 Purification of antibacterial compounds
Through silica gel column chromatography, antimicrobial compounds were purified, following the methodology explained [15]. As the eluting solvent methanol was employed, the column was packed with 60–120 mesh silica gel. The sample was loaded onto the column, and with methanol was eluted at a flow rate of one drop. Placed at the bottom of the column, the tubes were collected, representing the eluted fractions, and fractions considered most active were selected for further analysis.
The antibacterial compounds were analyzed using gas chromatography-mass spectrometry (GC-MS) against the NIST mass spectral library with spectral matching, a similarity threshold of greater than 85% to detect accurate compound assignment.
On an Agilent Technologies 6890-5973 N system equipped with a TG-5 ms Phenyl Methyl Siloxane capillary column (30 m × 250 μm × 0.25 μm), the analysis was performed. The mass detector operated in split mode, with helium gas serving as the carrier at a flow rate of 1.0 ml/min. The injector temperature was set to 230℃, and the oven temperature was initially set to 60℃ for 2 min, followed by a ramp of 10℃/min to reach 280℃ for an additional 8 min.
2.7 Using 96-well microtiter plates for antibacterial activity of active fractions
The active fractions obtained from Bacillus thermoamylovorans with antibacterial activity were evaluated using a 96-well microtiter plate assay, calculated by spectrophotometric analysis. This method allows for high-throughput screening of antimicrobial compounds against various bacterial strains. A 96-well microtiter plate was used for the assay, where each well contained a specific concentration of the active fractions from the purification process.
The microplate format allows for simultaneous testing of multiple samples, enhancing efficiency and reducing reagent use. Each well was inoculated with a standardized suspension of the target bacteria, such as uropathogenic strains. The inoculum size was adjusted to ensure consistent bacterial density across all wells. At 37℃, the plates were incubated for a predetermined period (typically 24 hours) to allow for bacterial growth and interaction with the antibacterial compounds. After incubation, the optical density (OD) of each well was measured at 600 nm (OD600) using a spectrophotometer. This measurement provides an indication of bacterial growth; lower OD values suggest higher antibacterial activity of the tested fractions.
2.8 Statistical analysis
The obtained data were subjected to a way analysis of variance (ANOVA) test to compare the means of various groups with each other. Results were expressed as mean ± standard Error (SE), and values of p > 0.05 were considered statistically non-significant, while p ≤ 0.05 and < 0.01,0.001 were considered significantly different, highly significantly different, respectively. The statistical analysis was carried out by SPSS (v 20).
3.1 Isolation and identification of strains
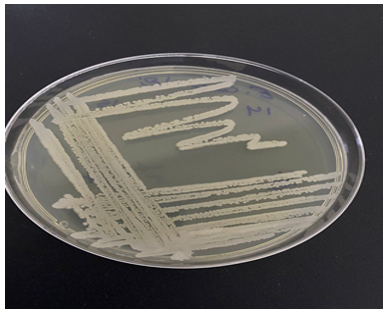
In this study, the isolation and identification of Bacillus spp. from skin samples were conducted to explore their potential for producing antimicrobial compounds. A total of 50 skin samples were analyzed, leading to the successful isolation of nine Bacillus strains. These strains were identified using Bergey’s Manual of Determinative Bacteriology, which relies on morphological so the colonies typically appear creamy white with circular in shape and the edges of the colonies are smooth, and they have a convex elevation on the tryptic soy agar (TSA) (Figure 2) and biochemical characteristics appear the isolate negative result for oxidase test and Positive for catalase test, as well as conventional techniques.
Figure 2. Growth of the Bacillus thermoamylovorans on optimal TSA medium
The isolated Bacillus strains were classified into three species: Bacillus subtilis (12%), Bacillus amyloliquefaciens (4%), and Bacillus thermoamylovorans (2%). The identification process was further validated using the VITEK 2 automated system, which provided rapid and accurate results that gave a 91% probability to diagnostic Bacillus thermoamylovorans (Table 2). This system is known for its high reliability in identifying both Gram-positive and Gram-negative bacteria, confirming the effectiveness of the methods employed. Each isolated strain was tested against uropathogenic bacteria to evaluate its antibacterial activity.
Table 2. VITEK 2 automated system for Bacillus thermoamylovorans
|
Biochemical Test |
Code |
Reaction (+/-) |
Biochemical Test |
Code |
Reaction (+/-) |
|
ProA |
8 |
(-) |
PheA |
7 |
(+) |
|
LeuA |
5 |
(-) |
AspA |
4 |
(-) |
|
LysA |
3 |
(-) |
BXYL |
1 |
(-) |
|
BNAG |
14 |
(-) |
TyrA |
13 |
(-) |
|
AlaA |
12 |
(-) |
AGAL |
11 |
(+) |
|
PyrA |
10 |
(+) |
BGAL |
9 |
(-) |
|
MdG |
24 |
(-) |
INO |
22 |
(-) |
|
GLYG |
21 |
(-) |
dGAL |
19 |
(-) |
|
CDEX |
18 |
(-) |
AppA |
15 |
(+) |
|
dMAN |
31 |
(-) |
GlyA |
30 |
(-) |
|
MTE |
29 |
(-) |
AMAN |
27 |
(-) |
|
MdX |
26 |
(-) |
ELLM |
25 |
(-) |
|
BGLU |
41 |
(-) |
IRHA |
39 |
(-) |
|
PLE |
37 |
(-) |
NAG |
36 |
(-) |
|
dMLZ |
34 |
(-) |
dMNE |
32 |
(+) |
|
dTRE |
48 |
(-) |
dTAG |
47 |
(-) |
|
AGLU |
46 |
(-) |
PVATE |
45 |
(-) |
|
PHC |
44 |
(-) |
BMAN |
43 |
(-) |
|
KAN |
59 |
(-) |
NaCl 6.5% |
58 |
(-) |
|
PSCNa |
56 |
(-) |
dRIB |
54 |
(+) |
|
dGLU |
53 |
(+) |
INU |
50 |
(-) |
|
POLYB1 |
63 |
(-) |
TTZ |
62 |
(-) |
|
ESC |
61 |
(-) |
OLD |
60 |
(-) |
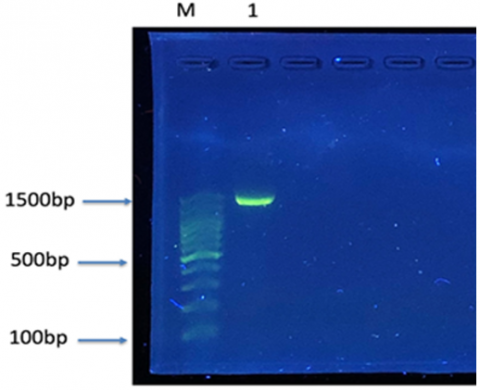
Figure 3. Identification of Bacillus thermoamylovorans by the 16S gene
The PCR product of 1500 bp was run alongside the DNA ladder on gel 1% agarose at 100V for 45 min. Visualized under UV light, the bands were: 1: Isolate of Bacillus thermoamylovorans, M=100 bp DNA ladder (transgen biotech) used as a DNA marker.
The isolation was also diagnosed using 16S rRNA sequencing, where this technique was used to accurately identify species through gene sequence analysis (Figure 3). In a previous study, 16S rRNA sequencing was utilized to identify different species of Bacillus, including Bacillus thermoamylovorans, which showed a genetic similarity of 93.9% with other species such as Bacillus thermolactis.
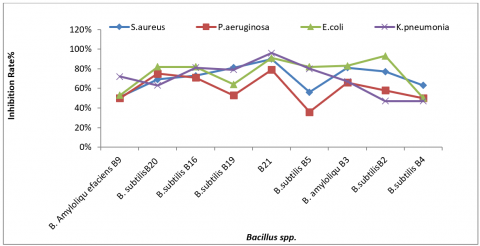
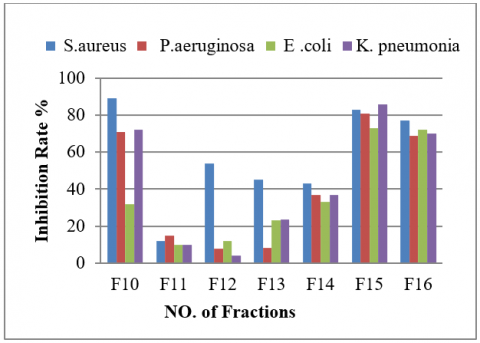
All Bacillus isolates demonstrated antibacterial properties; however, isolate B21 exhibited the most significant activity against uropathogenic strains except for the two isolates B16 and B4 (Table 3 and Figure 4).
Table 3. Antibacterial activity of Bacillus spp. (crude extract) on pathogenic bacteria
|
Bacillus spp. |
Type of Bacteria |
L.S.D. |
|||
|
S. aureus |
P. aeruginosa |
E. coli |
K. pneumonia |
||
|
B. amyloliquefaciens B9 |
51.66 ± 1.20 |
50.67 ± 0.67 |
53.33 ± 1.76 |
72.00 ± 1.52 |
8.02 * |
|
B. subtilis B20 |
69.00 ± 4.93 |
75.33 ± 2.91 |
82.33 ± 1.45 |
63.00 ± 2.08 |
8.94 * |
|
B. subtilis B16 |
73.33 ± 0.88 |
71.00 ± 1.00 |
82.33 ± 1.45 |
81.33 ± 1.76 |
7.63 * |
|
B. subtilis B19 |
81.00 ± 2.31 |
53.00 ± 1.53 |
64.33 ± 0.88 |
79.66 ± 1.20 |
9.36 * |
|
B. thermoamylovorans B21 |
90.66 ± 0.67 |
79.00 ± 2.08 |
91.66 ± 0.88 |
96.00 ± 1.00 |
7.69 * |
|
B. subtilis B5 |
56.33 ± 1.20 |
36.00 ± 2.08 |
82.33 ± 1.85 |
80.33 ± 2.73 |
10.77 * |
|
B. amyloliquefaciens B3 |
81.67 ± 2.33 |
66.00 ± 2.31 |
83.67 ± 1.76 |
67.33 ± 1.45 |
8.63 * |
|
B. subtilis B2 |
75.33 ± 1.20 |
57.66 ± 1.45 |
90.33 ± 1.45 |
47.00 ± 1.73 |
10.26 * |
|
B. subtilis B4 |
65.00 ± 1.00 |
51.67 ± 1.66 |
50.33 ± 2.60 |
48.33 ± 2.96 |
8.51 * |
|
L.S.D. |
11.37 * |
10.84 * |
10.79 * |
11.02 * |
--- |
|
* (P ≤ 0.05) |
|||||
Figure 4. Antibacterial activity of Bacillus spp. against tested microorganisms
3.2 Growth optimization of thermophilic bacteria, Bacillus thermoamylovorans
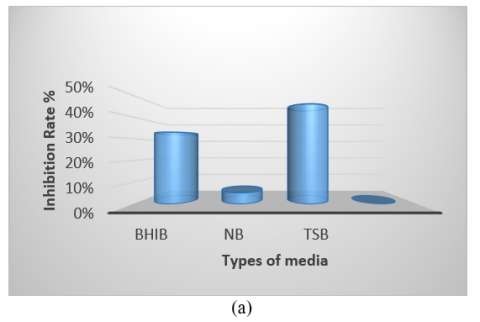
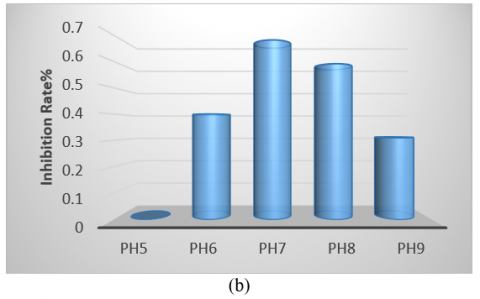
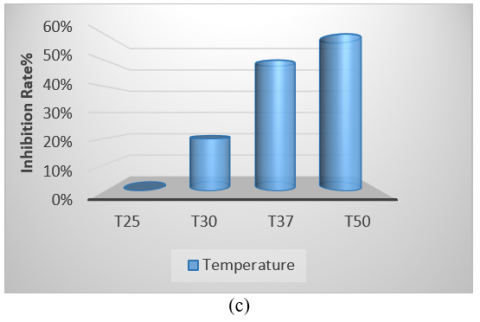
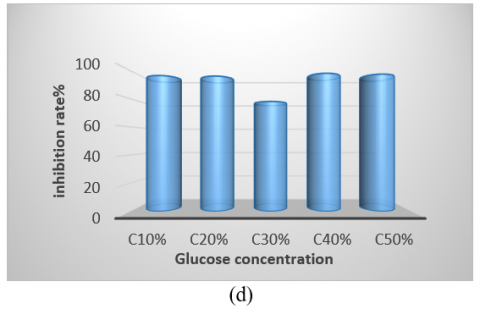
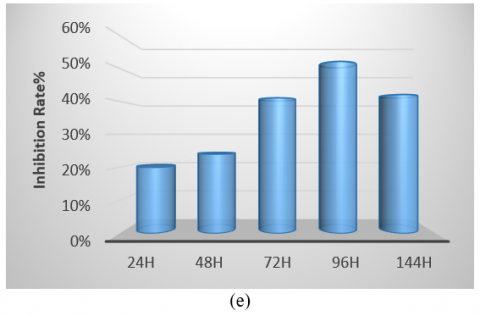
The bacterial isolate was cultivated in various culture media, showing the highest growth on tryptic soy broth and the lowest on Nutrient Broth. Therefore, identifying the optimal medium is crucial for enhancing antibacterial production (Figure 5(a)). Regarding pH, the optimum was found to be 7, as the initial pH of the medium is a significant factor in promoting microbial growth. Notably, the highest growth and production of antimicrobial compounds occurred at this pH compared to other values (Figure 5(b)). The isolate exhibited optimal growth at a temperature of 50℃, demonstrating its thermophile nature and ability to form spores, which facilitates the production of antibacterial compounds within the temperature range of 37-50℃. It was unable to grow at 30℃ (Figure 5(c)). Additionally, while the isolate grew at all glucose concentrations tested, the optimal concentration for carbon source utilization in antibacterial compound production was found to be 4% (Figure 5(d)). The period for producing pharmaceutical materials was determined to be 96 hours, which was more effective than other durations used in this study (Figure 5(e)).
Figure 5. (a) Different media affect the antibacterial activity of Bacillus thermoamylovorans; (b) Different pH values affect antibacterial activity; (c) Different temperatures affect activity; (d) Effect of different concentrations of glucose; (e) Antibacterial activity in various incubation periods
3.3 Extraction and purification of the active compounds
The strain was cultivated in tryptic soy broth at 50℃ with a 5% inoculum for four days, conditions that align with the optimal growth parameters for B. thermoamylovorans, this bacterium thrives at elevated temperatures, which enhances its metabolic activity and the production of bioactive compounds, including antimicrobial metabolites, while the filtration through filter paper Whatman No. 1 followed by 10,000 rpm centrifugation for 20 min effectively separated the cellular debris from the culture broth, allowing for the recovery of the supernatant that contains soluble metabolites.
This step is crucial for isolating compounds that may exhibit antibacterial activity, and the extraction method employed culture filtrate and ethyl acetate (1:1) in an equal volume, which is a common practice for isolating non-polar and semi-polar compounds. when the evaporation of the ethyl acetate layer at 50℃ yielded approximately 1.8 g of brown crude extract. The purification was conducted using column chromatography of silica gel, as described in the study [16]. Silica gel with a mesh size of 60–120 was utilized to pack the column, which is essential for effective separation of compounds due to its high surface area and porosity, and methanol was chosen as the eluting solvent.
The eluted fractions were collected in tubes placed at the bottom of the column, and subsequent analysis focused on identifying the most active fractions, which would likely contain the desired antimicrobial compounds. Following purification, the identified fractions were analyzed using GC-MS, specifically employing an Agilent Technologies 6890-5973 N system. This system is renowned for its sensitivity and accuracy in identifying volatile and semi-volatile compounds. The TG-5 ms Phenyl Methyl Siloxane capillary column (30 m × 250 μm × 0.25 μm) was used for separation, which provides excellent resolution for complex mixtures. After the purification process using a silica gel Column, 60 tubes of 5 ml each were collected, and their effectiveness against uropathogenic bacteria was tested after drying, then dissolved in DMSO (1%) to prepare a concentration of 500 µg/ml (Table 4 and Figure 6).
Figure 6. Antibacterial activity of fraction tubes against uropathogenic bacteria
Additionally, an analysis of the nature of the compounds in tubes 10-16 was conducted using GC-MS, which identified the nature of the antibiotic compounds, as detailed in Table 5.
Table 4. Antibacterial activity of fractions against test bacteria
|
Fraction (500 µg/ml) |
Type of Bacteria |
L.S.D. |
|||
|
S. aureus |
P. aeruginosa |
E. coli |
K. pneumonia |
||
|
F10 |
89.67 ± 0.88 |
71.00 ± 2.64 |
32.00 ± 1.52 |
72.33 ± 1.45 |
7.51 * |
|
F11 |
12.00 ± 1.52 |
15.00 ± 2.64 |
10.66 ± 0.67 |
10.66 ± 1.76 |
4.57 NS |
|
F12 |
54.33 ± 2.33 |
7.66 ± 1.45 |
12.33 ± 1.45 |
4.00 ± 2.08 |
11.38 * |
|
F13 |
45.67 ± 2.96 |
8.66 ± 1.85 |
23.66 ± 2.33 |
23.33 ± 0.88 |
9.77 * |
|
F14 |
43.67 ± 2.03 |
37.00 ± 2.51 |
33.00 ± 2.51 |
37.33 ± 4.05 |
7.41 * |
|
F15 |
83.33 ± 1.76 |
81.66 ± 1.67 |
73.33 ± 1.76 |
86.00 ± 2.31 |
7.92 * |
|
F16 |
77.00 ± 1.73 |
69.33 ± 2.40 |
72.33 ± 1.85 |
70.66 ± 1.20 |
6.98 * |
|
L.S.D. |
13.27 * |
14.55 * |
13.64 * |
16.55 * |
--- |
|
Values are means ±SD of triplicate measurements * (P ≤ 0.05), NS: Non-significant. |
|||||
Table 5. Active compounds identified in each fraction using GC-MS
|
Compound Name |
Area % |
Fraction |
Molecular Formula |
Molecular Weight (g/mol) |
Biological Importance |
|
9-Octadecenoic acid, (E) (Oleic acid) |
31.89 |
F10 |
C18H34O2 |
282.461 |
Cardioprotective, anti-inflammatory, antimicrobial, and skin health support. |
|
Cis-vaccenic acid |
27.88–35.15 |
F10-F16 |
C18H34O2 |
282.5 |
Anti-inflammatory, Anticancer, cardioprotective, improves immune response, lowers cholesterol. |
|
9,12-Octadecadienoic acid (Z,Z) (Linoleic acid) |
16.87 |
F10 |
C18H32O2 |
280.4 |
Antioxidant, anti-essential fatty acid, supports cell membrane integrity, anti-inflammatory, anticancer. |
|
Ascorbic acid (Vitamin C) |
14.06 |
F10, F11, F12 |
C6H8O6 |
176.12 |
Immune system support, Potent antioxidant, anticancer adjunct, tissue repair. |
|
1-Decanol, 5,9-dimethyl- |
13.38 |
F16 |
C12H26O |
186.33 |
Membrane disruption; antimicrobial potential, used industrially in coatings, solvents. |
|
Hexadecanoic acid, 2-hydroxyl-1-(hydroxymethyl) (Hydroxy palmitic acid) |
15.16 |
F11 |
C19H38O4 |
330.502 |
Skin moisturizer, antioxidant properties, and anti-inflammatory. |
|
Pentadecanoic acid |
12.26 |
F13 |
C15H30O2 |
242.40 |
Anticancer activities and anti-inflammatory. |
|
Dibutyl Phthalate |
10.84 |
F14 |
C16H22O4 |
278.34 |
Developmental effects and plasticizer, possible reproductive, environmental, and human health concerns. |
|
Diethyl Phthalate |
11.77 |
F14 |
C12H14O4 |
222.24 |
Endocrine disruption and reproductive effects. Used as a plasticizer; possibly, a potential environmental pollutant. |
|
(E)-9-Octadecenoic acid ethyl ester (Ethyl oleate) |
8.89 |
F10 |
C20H38O2 |
310.5 |
Enhances bioavailability of oleic acid, Anti-inflammatory, used in pharmaceutical formulations. |
|
Dodecanoic acid, 9-decen-1-yl ester (Decenoate ester) |
8.46 |
F10 |
C18H34O2 |
282.5 |
Membrane disruption in bacteria. Antimicrobial fatty acid ester. |
|
10,13-Octadecanoic acid, methyl ester |
7.53 |
F11 |
C19H34O2 |
294.5 |
Used in lipid formulations. Exhibits antibacterial effects. |
|
9-Eicosene, (E)- |
7.12 |
F14 |
C20H40 |
280.5 |
Minor bioactivity possible, pheromone in insects, used in chemical ecology studies. |
|
Tetradecanoic acid (Myristic acid) |
— |
F14 |
C14H28O2 |
228.37 |
Antifungal and Antibacterial, used in the food industry. and cosmetics. |
|
Tetradecane |
5.08 |
F12, F14 |
C14H30 |
198.39 |
Industrial applications: Used in solvents, with minor biological activity. |
|
Undecane |
5.47 |
F12 |
C11H24 |
156.31 |
Membrane interaction, hydrophobicity affects, and industrial solvent. |
|
Dodecane |
5.84 |
F11, F12 |
C12H26 |
170.33 |
Acts as a solvent, mild antimicrobial effects/membrane fluidizer. |
The successful isolation of Bacillus spp. was confirmed using advanced identification techniques, including the VITEK 2 system and 16S rRNA sequencing, alongside morphological and biochemical analyses (Table 2) [16]. This multi-level approach provided accurate identification of the isolate and ensured the reliability of subsequent experimental results.
Notably, Bacillus thermoamylovorans exhibited a broader antibacterial spectrum against Gram-negative than Gram-positive bacteria. This may be attributed to structural differences, as Gram-negative bacteria possess an outer membrane enriched with lipopolysaccharides (LPS), which act as an additional barrier limiting the entry of antimicrobial agents. To further clarify these interactions, future studies should examine the molecular binding of fatty acids such as cis-vaccenic acid with bacterial LPS. Understanding these mechanisms could contribute to optimizing the therapeutic potential of natural antimicrobials.
The findings are consistent with previous reports indicating that Bacillus species synthesize a wide range of antimicrobial metabolites, including lipopeptides, fatty acids, and bacteriocins, with broad inhibitory effects on pathogenic bacteria [17]. These compounds act through multiple mechanisms, such as disruption of membrane integrity, inhibition of peptidoglycan biosynthesis, interference with bacterial metabolism, induction of oxidative stress via ROS production, and inhibition of DNA replication and repair.
Cultivation of the isolate under optimized conditions promoted the production of bioactive metabolites, as reported in earlier studies [18]. Ethyl acetate proved effective in extracting secondary metabolites, particularly phenolic acids and flavonoids, which are known for their antimicrobial activities [19]. Partial purification using silica gel column chromatography allowed the separation of these compounds, and fractions 10–16 exhibited strong antibacterial activity, with the highest effects recorded in tubes 13–16. The variation in antibacterial activity among the fractions likely reflects differences in compound concentrations and structural characteristics, which influence their ability to interact with bacterial cell walls and intracellular targets [20, 21].
Further analysis of the active fractions using GC–MS identified diverse bioactive metabolites (Table 5). The major constituents included unsaturated fatty acids such as oleic acid, vaccenic acid, and linoleic acid, which are well documented for their antimicrobial, anti-inflammatory, antioxidant, and cardioprotective properties. Their main mode of action involves disrupting microbial membranes and increasing permeability, leading to cell lysis. However, their clinical application is limited by poor aqueous solubility and environmental instability; thus, novel delivery systems such as nanoemulsions and liposomes have been proposed to enhance their therapeutic potential [22-24].
Saturated fatty acids (e.g., pentadecanoic acid, myristic acid) were also identified, displaying antibacterial and antifungal properties. At higher concentrations, however, they may interfere with host lipid homeostasis, which restricts their practical application [25]. In addition, fatty acid esters such as ethyl oleate and decenoate ester were detected; these compounds are important for enhancing the bioavailability and stability of their parent fatty acids, and they have been incorporated into pharmaceutical formulations for improved therapeutic outcomes.
Another important metabolite, ascorbic acid (vitamin C), was detected in several fractions and is known for its antioxidant capacity, immune modulation, and synergistic effects with antimicrobial agents [26]. Conversely, compounds such as dibutyl phthalate and diethyl phthalate were considered undesirable contaminants, given their endocrine-disrupting and reproductive toxicity effects [27]. Likewise, alkanes (tetradecane, undecane, dodecane) exhibited only minor biological activity, mainly acting as solvents or metabolic byproducts.
Among all identified compounds, cis-vaccenic acid was particularly significant, constituting up to 66% of the composition in fraction 15. This fatty acid showed pronounced antibacterial activity, especially against urinary tract infection–associated pathogens. Its mechanism is multifaceted, involving direct membrane disruption, inhibition of biofilm formation, enhancement of antibiotic efficacy, and interference with bacterial quorum sensing [27-29]. These combined activities highlight cis-vaccenic acid as a promising natural antimicrobial candidate for further research.
In summary, the results demonstrate that Bacillus thermoamylovorans produces a diverse range of bioactive compounds, with fatty acids representing the dominant antimicrobial metabolites. Their broad mechanisms of action, natural abundance, and relative safety emphasize their potential in developing alternative therapies against multidrug-resistant pathogens. However, challenges such as solubility, stability, and cytotoxicity must be addressed through formulation strategies and further mechanistic studies to realize their clinical and industrial applications.
This study highlights Bacillus thermoamylovorans as a promising source of novel bioactive compounds, derivatives of fatty acids, for combating antibiotic-resistant pathogens. By these compounds the potent antibacterial activities demonstrated, combined with lack of cross-resistance, their novel structural profiles and emphasize their potential utility in the development to address the global challenge of antibiotic resistance of new therapeutic agents. An important foundation this work lays for future mechanism of action studies, biochemical characterization, and potential pharmaceutical applications.
The special thanks from the authors to Mustansiriyah University, College of Science, and Department of Biology, Baghdad, Iraq.
[1] Lineback, C.B., Nkemngong, C.A., Wu, S.T., Li, X., Teska, P.J., Oliver, H.F. (2018). Hydrogen peroxide and sodium hypochlorite disinfectants are more effective against Staphylococcus aureus and Pseudomonas aeruginosa biofilms than quaternary ammonium compounds. Antimicrobial Resistance & Infection Control, 7(1): 154.
[2] Edwar, D.A., Naji, I.N., Aboul-Ela, H.M. (2023). Investigation the Role of various antiseptics on the prevalence of skin microbiota and post cesarean surgery infections. Al-Mustansiriyah Journal of Science, 34(3): 1-9. https://doi.org/10.23851/mjs.v34i3.1319
[3] Grice, E.A., Segre, J.A. (2011). The skin microbiome. Nature Reviews Microbiology, 9(4): 244-253. https://doi.org/10.1038/nrmicro2537
[4] Blaskovich, M.A., Elliott, A.G., Kavanagh, A.M., Ramu, S., Cooper, M.A. (2019). In vitro antimicrobial activity of acne drugs against skin-associated bacteria. Scientific Reports, 9(1): 14658. https://doi.org/10.1038/s41598-019-50746-4
[5] Ajuna, H.B., Lim, H.I., Moon, J.H., Won, S.J., Choub, V., Choi, S.I., Yun, J.Y., Ahn, Y.S. (2023). The prospect of hydrolytic enzymes from Bacillus species in the biological control of pests and diseases in forest and fruit tree production. International Journal of Molecular Sciences, 24(23): 16889. https://doi.org/10.3390/ijms242316889
[6] Simandjuntak, S., Samuel, M.Y. (2018). Isolation and identification of thermophilic bacteria, producer of amylase enzyme, from Lake Linow, North Sulawesi. Journal of Pure and Applied Microbiology, 12(2): 543-554. https://doi.org/10.22207/JPAM.12.2.13
[7] Dashti, A.A., Jadaon, M.M., Abdulsamad, A.M., Dashti, H.M. (2009). Heat treatment of bacteria: A simple method of DNA extraction for molecular techniques. Kuwait Medical Journal, 41(2): 117-122.
[8] Ligozzi, M., Bernini, C., Bonora, M.G., De Fatima, M., Zuliani, J., Fontana, R. (2002). Evaluation of the VITEK 2 system for identification and antimicrobial susceptibility testing of medically relevant gram-positive cocci. Journal of Clinical Microbiology, 40(5): 1681-1686. https://doi.org/10.1128/jcm.40.5.1681-1686.2002
[9] Tran, C., Cock, I.E., Chen, X., Feng, Y. (2022). Antimicrobial Bacillus: metabolites and their mode of action. Antibiotics, 11(1): 88. https://doi.org/10.3390/antibiotics11010088
[10] Mohammed, N.S., Hassan, M.J.M., Mahdi, A.S. (2019). New spectrophotometric method for determination of ceftazidime in pure form and pharmaceutical dosages. Al-Mustansiriyah Journal of Science, 30(3): 47-52. http://doi.org/10.23851/mjs.v30i3.661
[11] Geraldi, A., Famunghui, M., Abigail, M., Siona Saragih, C.F., et al. (2022). Screening of antibacterial activities of Bacillus spp. isolated from the Parangkusumo coastal sand dunes, Indonesia. BIO Integration, 3(3): 132-137. http://doi.org/10.15212/bioi-2022-0005
[12] Chen, L., Wang, C., Su, J. (2023). Understanding the effect of different glucose concentrations in the oligotrophic bacterium Bacillus subtilis BS-G1 through transcriptomics analysis. Microorganisms, 11(10): 2401. https://doi.org/10.3390/microorganisms11102401
[13] Wilks, J.C., Kitko, R.D., Cleeton, S.H., Lee, G.E., Ugwu, C.S., Jones, B.D., BonDurant, S.S., Slonczewski, J.L. (2009). Acid and base stress and transcriptomic responses in Bacillus subtilis. Applied and Environmental Microbiology, 75(4): 981-990. https://doi.org/10.1128/AEM.01652-08
[14] Aqel, H., Farah, H., Al-Hunaiti, A. (2024). Ecological versatility and biotechnological promise: Comprehensive characterization of the isolated thermophilic Bacillus strains. Plos One, 19(4): e0297217. https://doi.org/10.1371/journal.pone.0297217
[15] Dhaini, H.K., Khalil, M.I., Hajj, R.E. (2025). The antimicrobial potential of actinomycetes isolated from marine soils in Tyre City Beach, Lebanon: A promising source of novel bioactive metabolites. Applied Microbiology, 5(1): 27. https://doi.org/10.3390/applmicrobiol5010027
[16] Thapa, A., Budhathoki, A., Sapkota, A., Sainju, M., Shrestha, P., Pant, S.P. (2021). Isolation, identification and screening of Bacillus species with antimicrobial activity from different soil samples of Kathmandu Valley. Nepal Journal of Biotechnology, 9(2): 1-6.
[17] Caulier, S., Nannan, C., Gillis, A., Licciardi, F., Bragard, C., Mahillon, J. (2019). Overview of the antimicrobial compounds produced by members of the Bacillus subtilis group. Frontiers in Microbiology, 10: 302. https://doi.org/10.3389/fmicb.2019.00302
[18] Sehim, A.E., Amin, B.H., Yosri, M., Salama, H.M., Alkhalifah, D.H., Alwaili, M.A., Abd Elghaffar, R.Y. (2023). GC-MS analysis, antibacterial, and anticancer activities of Hibiscus sabdariffa L. methanolic extract: In vitro and in silico studies. Microorganisms, 11(6): 1601. https://doi.org/10.3390/microorganisms11061601
[19] Jadimurthy, R., Jagadish, S., Nayak, S.C., Kumar, S., Mohan, C.D., Rangappa, K.S. (2023). Phytochemicals as invaluable sources of potent antimicrobial agents to combat antibiotic resistance. Life, 13(4): 948. https://doi.org/10.3390/life13040948
[20] Singh, S.P., Qureshi, A., Hassan, W. (2021). Mechanisms of action by antimicrobial agents: A review. McGill Journal of Medicine, 19(1). https://doi.org/10.26443/mjm.v19i1.217
[21] Desbois, A.P., Smith, V.J. (2010). Antibacterial free fatty acids: Activities, mechanisms of action and biotechnological potential. Applied Microbiology and Biotechnology, 85(6): 1629-1642. https://doi.org/10.1007/s00253-009-2355-3
[22] Yoon, B.K., Jackman, J.A., Valle-González, E.R., Cho, N.J. (2018). Antibacterial free fatty acids and monoglycerides: biological activities, experimental testing, and therapeutic applications. International Journal of Molecular Sciences, 19(4): 1114. https://doi.org/10.3390/ijms19041114
[23] Salvia-Trujillo, L., Qian, C., Martín-Belloso, O., McClements, D.J. (2013). Influence of particle size on lipid digestion and β-carotene bioaccessibility in emulsions and nanoemulsions. Food Chemistry, 141(2): 1472-1480. https://doi.org/10.1016/j.foodchem.2013.03.050
[24] Calder, P.C. (2009). Fatty acids and immune function: Relevance to inflammatory bowel diseases. International Reviews of Immunology, 28(6): 506-534. https://doi.org/10.3109/08830180903197480
[25] Hassuna, N.A., Rabie, E.M., Mahd, W.K.M., Refaie, M.M., Yousef, R.K.M., Abdelraheem, W.M. (2023). Antibacterial effect of vitamin C against uropathogenic E. coli in vitro and in vivo. BMC Microbiology, 23(1): 112. https://doi.org/10.1186/s12866-023-02856-3
[26] Tran, H.T., Lin, C., Bui, X.T., Nguyen, M.K., et al. (2022). Phthalates in the environment: characteristics, fate and transport, and advanced wastewater treatment technologies. Bioresource Technology, 344: 126249. https://doi.org/10.1016/j.biortech.2021.126249
[27] Kumar, P., Kumar, D., Pal, S., Singh, S. (2025). Plant secondary metabolites in defense against phytopathogens: Mechanisms, biosynthesis, and applications. Physiological and Molecular Plant Pathology, 138: 102639. https://doi.org/10.1016/j.pmpp.2025.102639
[28] Casillas-Vargas, G., Ocasio-Malavé, C., Medina, S., Morales-Guzmán, C., Del Valle, R.G., Carballeira, N.M., Sanabria-Ríos, D.J. (2021). Antibacterial fatty acids: An update of possible mechanisms of action and implications in the development of the next-generation of antibacterial agents. Progress in Lipid Research, 82: 101093. https://doi.org/10.1016/j.plipres.2021.101093
[29] Karuppiah, V., Seralathan, M. (2022). Quorum sensing inhibitory potential of vaccenic acid against Chromobacterium violaceum and methicillin-resistant Staphylococcus aureus. World Journal of Microbiology and Biotechnology, 38(8): 146. https://doi.org/10.1007/s11274-022-03335-z