Bagus Prasetyo Aji*![]() | Fuad
| Fuad![]() | Wahyu Isroni
| Wahyu Isroni![]() | Lisa Nur Hidayah
| Lisa Nur Hidayah![]() | Adi Tiya Yanuar
| Adi Tiya Yanuar![]() | Anjas Sasana Bahari
| Anjas Sasana Bahari![]()
© 2025 The authors. This article is published by IIETA and is licensed under the CC BY 4.0 license (http://creativecommons.org/licenses/by/4.0/).
OPEN ACCESS
Mini trawl fisheries remain widely used in Indonesian coastal waters despite regulatory bans on trawl operations, raising ecological concerns due to their low selectivity. This study analyzed 125 mini trawl landings in Lamongan, East Java, recording 8,105 kg of biomass across 32 species. The catch was dominated by Stolephorus sp. (51.4%) and Leiognathus splendens (21.8%). Diversity indices indicated low richness (R₁ = 2.06), low evenness (E = 0.16), and high dominance (C = 1.00), indicating ecological simplification due to intensive fishing pressure. One-way ANOVA showed significant differences in mean catch weight among species (F = 14.63; p < 0.01), although most demersal and benthic taxa exhibited low and statistically similar weights. Hierarchical Cluster Analysis (HCA) revealed that species with distinct morphologies—such as crustaceans and cephalopods—clustered alongside fishes, confirming gear non-selectivity. These results underscore the need for morphology-based gear modifications, including square mesh codends and bycatch reduction devices (BRDs), to reduce ecological impacts. Integrating biological traits, species grouping, and statistical evidence provides a basis for ecosystem-based fisheries management in tropical coastal systems.
mini-trawl, catch composition, species diversity, community structure, morphological clustering
Capture fisheries are a key sector of the global economy, food security, and livelihood for coastal communities, particularly in maritime countries such as Indonesia [1-3]. This sector contributes significantly to regional and national economic growth, animal protein supply, and employment. However, the ecological sustainability of fishing practices is under threat, primarily due to the use of non-selective and environmentally damaging fishing gear [4, 5]. In tropical marine ecosystems with high biodiversity and ecological sensitivity, such fishing gear is causing damage to habitats and species communities, exacerbating the sustainability concerns.
Trawling is widely recognised as one of the most destructive fishing methods due to its low selectivity and broad sweeping capacity [6, 7]. It captures target and non-target species, including demersal fish, molluscs, crustaceans, and juvenile organisms. Numerous studies have documented its impacts, such as the degradation of seafloor habitats, disruption of benthic communities, reduced resilience of fish populations, and acceleration of carbon release from sediments [8-10]. High bycatch rates and discards of low-value species exacerbate ecological stress and waste biological resources [11, 12].
Despite the national ban on trawling under Ministerial Regulation No. 2 of 2015 [13], mini trawl operations remain prevalent in Indonesian coastal regions, including Lamongan [14, 15]. Socioeconomic dependency and enforcement challenges have hindered regulation implementation. Local fishers often perceive Mini trawls as efficient but have been linked to habitat degradation, inter-fisher conflicts, and indiscriminate capture of ecologically important species. However, scientific studies on their ecological impacts remain limited, especially at the local level.
Despite the widespread use of mini-trawls in tropical small-scale fisheries, limited information is available on how this gear influences community composition, biodiversity patterns, and morphological selectivity. Previous studies have primarily focused on biomass yields and economic contributions, whereas ecological implications such as species dominance, recruitment overfishing, and morphological vulnerability have remained insufficiently quantified. Addressing these knowledge gaps is essential for evaluating mini-trawl fisheries' ecological sustainability and informing management interventions that integrate biological and ecosystem-based perspectives.
This study hypothesized that mini-trawl fisheries exhibit low selectivity across morphological groups, resulting in a catch structure dominated by small pelagic species and undersized individuals, with implications for community organization and ecosystem resilience.
Specifically, the objectives of this research were to: describe the catch composition of mini-trawl fisheries in Lamongan waters; analyze community structure, including species richness, diversity, evenness, and dominance indices; and assess morphological clustering of captured species to evaluate patterns of selectivity.
2.1 Study area
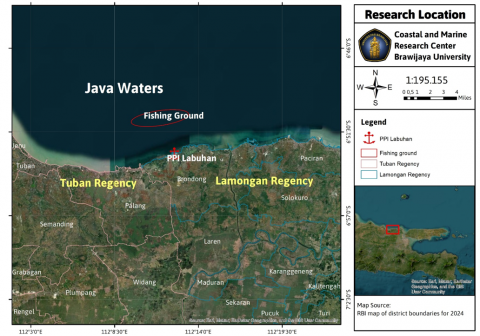
The research was conducted from March to April 2024 at the Labuhan Village Fish Auction Site, Lamongan Regency, East Java, Indonesia (6°52'57.49" S 112°12'16.83" E) (Figure 1). This location was chosen because it is one of the leading centres for fishing activities using mini trawl nets, which have high operational intensity and serve as the primary landing point for local fishermen's catches. The fishing area is located in the coastal waters of Lamongan District, with a depth of 15–20 metres.
Figure 1. Research location
2.2 Data collection
A total of 125 vessels were randomly selected during the observation period. All sampling was carried out within the same fishing season at a single landing site to minimize temporal variation. Gear size and configurations (net length, mesh size, and codend dimensions) were verified through fisher interviews and dockside inspection to ensure comparability across vessels.
Fish identification was performed based on external morphological features such as body shape, total length, fin arrangement, coloration, and tail morphology. Representative samples (1-5 individuals per species) were collected and identified to species level using standard references [16-20]. Identification was validated using expert cross-checks and online databases such as FishBase.
The study employed direct observation and semi-structured interviews with fishers to obtain data on vessel specifications, fishing gear characteristics, and fishing locations. Random sampling was used to select vessels' landing catches during the observation period.
2.3 Data analysis
Catch composition was quantified as the proportion of each species' biomass relative to total catch weight. The number of individuals was estimated using a subsampling approach. For each species, 1-5 representative samples were weighed to calculate the mean individual weight. This mean weight was then divided by the total biomass of each species to estimate the total number of individuals. This method was applied to account for the dominance of small-bodied pelagic taxa that occurred in very high abundance. Community indices included Shannon-Wiener diversity (H’), Pielou’s evenness (E), Odum’s dominance index (C), and Margalef’s richness index (R1).
Hierarchical Cluster Analysis (HCA) was used to classify species based on morphological similarity. The input variable included body shape, fin position, and total length, which were selected because they influence gear retention and mesh escapement. Euclidean distance and the UPGMA method were applied to construct a species cluster.
A one-way Analysis of Variance (ANOVA) was applied to evaluate differences in mean biomass among species. Before analysis, the assumptions of normality and homogeneity of variance were tested using the Shapiro–Wilk and Levene's tests, respectively. As both assumptions were satisfied (p > 0.05), the dataset was deemed appropriate for parametric analysis. Where significant effects were detected (p < 0.05), Tukey's Honestly Significant Difference (HSD) test was used to identify pairwise differences between species. All analyses used IBM SPSS Statistics version 26.0 [21, 22].
3.1 Catch composition
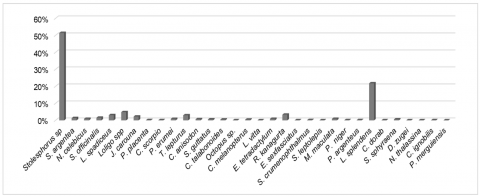
The observation (Table 1) revealed that the total biomass of fish and marine organisms caught amounted to 8,105 kg, comprising an estimated 3,416,234 individuals from 32 distinct species. The catch composition was dominated by Stolephorus sp. (anchovy) at 51.42%, Leiognathus splendens (ponyfish) at 21.79% and Loligo spp at 4.83%. Collectively, these three species accounted for over 70% of the total catch weight, indicating a pronounced dominance in the structure of the fish community (Figure 2).
Table 1. Catch composition
|
No. |
Species |
Fish Name |
Weight (Kg) |
Taxonomy |
Ecological Habitat |
|
1 |
Stolesphorus sp. |
Anchovy |
4,167.72 |
O |
Pelagic |
|
2 |
Leiognathus splendens |
Ponyfish |
1,766.49 |
O |
Demersal |
|
3 |
Loligo spp. |
Squid |
391.63 |
M |
Demersal |
|
4 |
Rastrelliger kanagurta |
Long-jawed Mackerel |
271.39 |
O |
Pelagic |
|
5 |
Lagocephalus spadiceus |
Pufferfish |
248.76 |
O |
Demersal |
|
6 |
Trichiurus lepturus |
Cutlassfish |
241.23 |
O |
Pelagic |
|
7 |
Sepia officinalis |
Common cuttlefish |
124.81 |
M |
Demersal |
|
8 |
Johnius carouna |
Croaker fish |
181.67 |
O |
Demersal |
|
9 |
Saurida argentea |
Shortin saury |
109.36 |
O |
Demersal |
|
10 |
Nemipterus celebicus |
Celebes Threadfin Bream |
72.49 |
O |
Pelagic |
|
11 |
Mene maculata |
Moonfish |
72.40 |
O |
Demersal |
|
12 |
Psettodes erumei |
Flatfish |
70.80 |
O |
Demersal |
|
13 |
Eleutheronema tetradactylum |
Giant threadfin |
71.51 |
O |
Demersal |
|
14 |
Charybdis anisodon |
Oceanic rowing crab |
53.38 |
Cr |
Demersal |
|
15 |
Scomberomorus guttatus |
Spanish mackerel |
46.21 |
O |
Pelagic |
|
16 |
Sphyraena sphyraena |
Barracuda |
46.01 |
O |
Demersal |
|
17 |
Congresox talabonoides |
Snake eel |
38.37 |
O |
Demersal |
|
18 |
Carcharhinus melanopterus |
Shark |
34.70 |
Ch |
Demersal |
|
19 |
Selar crumenophthalmus |
Bigeye scad |
12.80 |
O |
Pelagic |
|
20 |
Pampus argenteus |
Silver pomfret |
12.65 |
O |
Demersal |
|
21 |
Placuna placenta |
Windowpane oyster |
10.86 |
M |
Demersal |
|
22 |
Cloridopsis scorpio |
Mantis Shrimp |
9.71 |
Cr |
Demersal |
|
23 |
Parastromateus niger |
Black pomfret |
9.30 |
O |
Demersal |
|
24 |
Caranx ignobilis |
Travelly |
8.01 |
O |
Pelagic |
|
25 |
Chirocentrus dorab |
Dorab wolf-herring |
5.85 |
O |
Demersal |
|
26 |
Octopus sp. |
Octopus |
5.60 |
M |
Demersal |
|
27 |
Epinephelus sexfasciatus |
Grouper |
5.55 |
O |
Demersal |
|
28 |
Selaroide leptolepis |
Yellowstrip scad |
4.50 |
O |
Pelagic |
|
29 |
Netuma thalassina |
Ariid catfish |
4.20 |
O |
Demersal |
|
30 |
Lutjanus vitta |
Brownstrip red snapper |
3.10 |
O |
Demersal |
|
31 |
Dasyatis zugei |
Stingray |
3.10 |
Ch |
Demersal |
|
32 |
Penaeus merguiensis |
Shrimp |
1.17 |
Cr |
Demersal |
Figure 2. Catch composition in Lamongan mini trawl fisheries
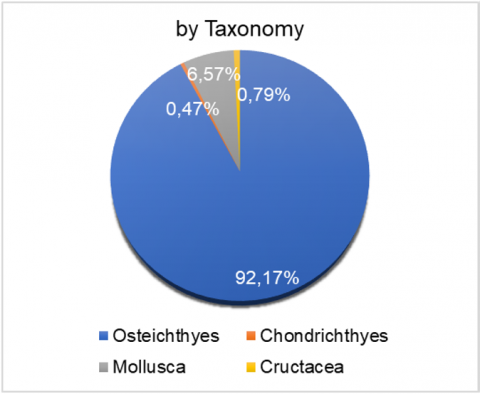
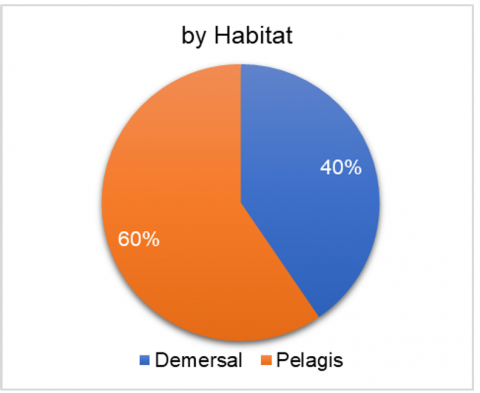
Taxonomically (Figure 3(a)), the catch was dominated by the Osteichthyes group, comprising 92.17% of the total individuals. Mollusca accounted for 6.75%, followed by Chondrichthyes (Charcharhinus melanopterus and Dasyatis zugei) at 0.79%, and Crustacea at 0.47%. In terms of ecological habitat, 60% of the captured species were demersal, while the remaining 40% were pelagic.
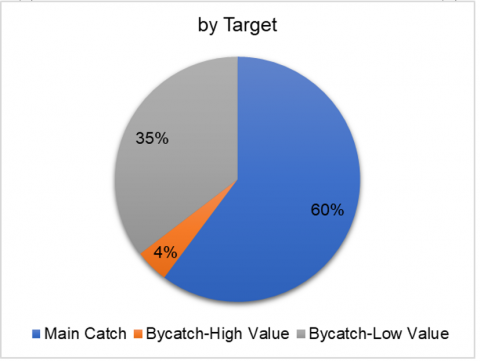
From an economic perspective (Figure 3(b)), approximately 60% of the catch consisted of high-value target species, including Stolephorus sp., Leiognathus splendens, and Rastrelliger kanagurta; 4% comprised high-value bycatch species such as Sphyraena sphyraena and Epinephelus sexfasciatus; while the remaining 35% were categorized as low-value bycatch (Figure 3(c)). This composition reflects the dominance of small pelagic and demersal species that typically form schooling aggregations, along with a relatively broad taxonomic diversity.
Figure 3. Proportion of catch based on (a) taxonomy; (b) ecological habitat; (c) catch target
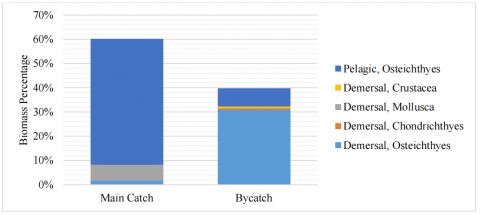
Figure 4 illustrates the relative biomass composition of target and bycatch species, categorized based on habitat (pelagic and demersal) and major taxonomic groups (Osteichthyes, Crustacea, Mollusca, and Chondrichthyes). The target catch was predominantly composed of pelagic, Osteichthyes, which accounted for over 51% of the total biomass. This group was mainly represented by small schooling species such as Stolephorus sp. and Rastrelliger kanagurta. A smaller fraction of demersal Mollusca, including Loligo sp. and Sepia officinalis, was also recorded, suggesting a degree of vertical overlap in gear operation that extends into both pelagic and demersal strata.
3.2 Species diversity index
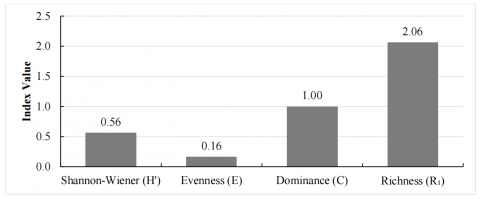
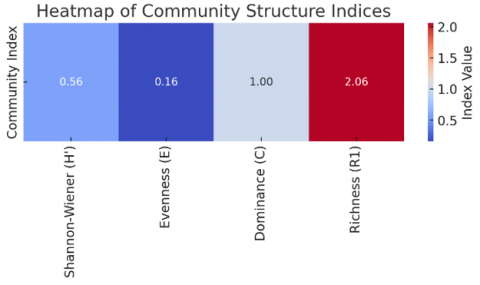
Based on the calculated indices (Table 2), the fish community structure in Lamongan waters exhibited low species diversity (H′ = 0.56), very low evenness (E = 0.16), and complete dominance by a single taxon (C = 1.00), indicating a highly imbalanced assemblage. Species richness (R₁ = 2.06) was also low, suggesting limited taxonomic representation within the catch. These results are further visualized in Figure 5 (bar plot) and Figure 6 (heatmap), emphasizing the disparity between diversity components.
The bar plot displays the absolute index values, while the heatmap accentuates the contrast in community metrics through color gradients. Together, these visualizations support the interpretation that the mini trawl fishery exerts high pressure on the ecosystem, promoting opportunistic dominance (e.g., Stolephorus sp.) and reducing overall ecological complexity. Such structural patterns are commonly associated with the long-term use of non-selective fishing gear in tropical multispecies fisheries.
Table 2. Community structure index value
|
Parameter |
Index Value |
|
Number of species (S) |
32 |
|
Number of individuals (N) |
3,416,234 |
|
Diversity indices (H’) |
0.56 |
|
Evenness indices (E) |
0.16 |
|
Dominan indeces (C) |
1 |
|
Species richness indeces (R1) |
2.06 |
Figure 5. Bar plot of community structure indices showing low diversity (H′ = 0.56), low evenness (E = 0.16), high dominance (C = 1.00), and low richness (R1 = 2.06)
Figure 6. Heatmap of community structure indices illustrating index value gradients, indicating an ecologically simplified fish community
3.3 Testing variations in catch weight between species
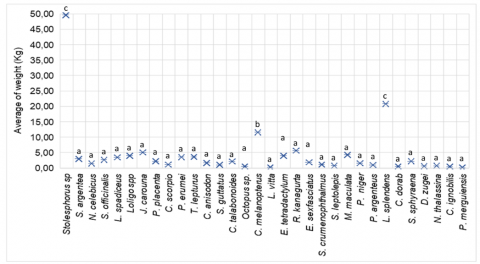
One-way Analysis of Variance (ANOVA) showed a statistically significant difference in mean catch biomass among the five dominant species (F = 14.63; p < 0.001; Figure 7). The post-hoc Tukey's Honestly Significant Difference (HSD) test revealed that Stolephorus sp. (33.34 ± 5.71 kg) differed significantly from all other species (p < 0.01), followed by Leiognathus splendens (14.13 ± 2.30 kg), which also showed significant differences from Loligo sp. (3.13 ± 1.05 kg), Penaeus merguiensis (0.009 ± 0.002 kg), and Placuna placenta (0.087 ± 0.015 kg). No significant difference was observed between Penaeus merguiensis and Placuna placenta (p > 0.05). As visualized in Figure 6, the biomass distribution was skewed toward pelagic species, with clear separation indicated by distinct statistical groupings.
Figure 7. Mean catch weight per species (kg/unit) based on one-way ANOVA with Tukey's post hoc test (p < 0.05)
3.4 Hierarchical Cluster Analysis (HCA)
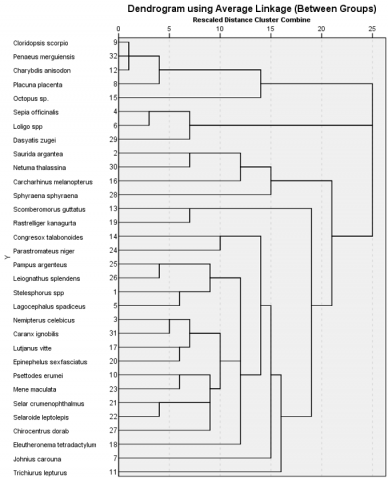
Species relationships were analysed using Hierarchical Cluster Analysis (HCA) (Figure 8) with an average linkage method, resulting in a dendrogram delineating several major clusters. Based on morphological similarities, the captured species were grouped into clusters exhibiting varying levels of taxonomic distance. Species with comparable body forms and tail morphologies, such as Sepia officinalis, Loligo sp., and Octopus sp., were grouped at lower similarity distances. In contrast, Cloridopsis scorpio, Penaeus merguiensis, and Placuna placenta formed distinct clusters, reflecting greater morphological divergence from the Osteichthyes group.
Figure 8. Dendrogram of Hierarchical Cluster Analysis (HCA) showing species groupings based on morphological
4.1 Catch composition
The composition of mini-trawl catches in Lamongan waters was dominated by small pelagic and small to medium-sized demersal fishes, reflecting the gear's low selectivity and broad sweeping capacity. The predominance of Stolephorus sp. and Leiognathus splendens can be explained by several technical and ecological factors. Mini-trawls are typically operated in shallow coastal waters where small pelagic fishes naturally concentrate. The small mesh size in the codend reduces gear selectivity, allowing juveniles and undersized individuals to be retained [23, 24]. In addition, the frequent one-day fishing operations increase exploitation pressure in nearshore ecosystems. These operational characteristics collectively explain the skewed species composition, where small pelagic taxa dominate while larger demersal and benthic organisms are underrepresented. Such outcomes highlight how gear design and fishing practice strongly shape ecological patterns in mini-trawl fisheries.
The catch was dominated by bony fishes (Osteichthyes), accounting for 92.17% of the total individuals, followed by Mollusca (6.57%), Chondrichthyes (0.79%), and Crustacea (0.47%). Although these non-target taxa occurred in small proportions, their capture indicates a significant bycatch potential, particularly for groups with ecological or conservation value such as cephalopods, sharks, and rays [8, 11, 25]. From a habitat perspective, 60% of the species were demersal and 40% pelagic. While mini-trawls are categorized as bottom trawls, their configuration and towing dynamics allow the nets to sweep demersal and pelagic strata simultaneously. This overlap explains why pelagic schooling species dominate the catch while demersal invertebrates and benthic fishes are also consistently present. Such cross-strata exploitation not only broadens the ecological footprint of mini-trawls but also exacerbates conflicts between efficiency (high pelagic biomass) and sustainability (incidental capture of ecologically sensitive taxa) [9, 26].
Classification by economic value indicated that only 60% of the catch consisted of high-value target species, 35% was low-value bycatch, and 4% comprised high-value non-target species. This composition illustrates economic benefits and inefficiencies: while pelagic schooling fishes such as Stolephorus sp. provide substantial biomass, the incidental capture of low-value or juvenile fishes represents a loss of potential future yield. Retaining juveniles of commercially important species suggests hidden economic costs by reducing recruitment into the fishery [27, 28]. Moreover, the disproportionate capture of small, short-lived taxa reflects the "fishing down the food web" phenomenon [29], where lower trophic levels replace top predators due to gear selectivity and stock depletion. Such outcomes emphasize that mini-trawls generate immediate catch efficiency but at the expense of long-term sustainability, underscoring the trade-off between short-term economic gain and the ecological resilience of tropical coastal fisheries [23].
The predominance of small-bodied, fast-growing species such as Stolephorus sp. and Leiognathus splendens in the catch composition reflects the strong selectivity of mini-trawl gear toward schooling pelagic taxa inhabiting surface and subsurface strata. These species are classified as r-strategists, meaning they can adapt ecologically to disturbance. However, they are also vulnerable to continuous fishing pressure due to their short life cycles and high reproduction rates [24]. The observed catch structure is consistent with the "fishing down the food web" hypothesis, where lower trophic-level species gradually replace long-lived top predators as a result of overexploitation and gear-induced selectivity [9, 29].
Such shifts in trophic structure have been widely documented in tropical multispecies trawl fisheries, particularly those lacking spatial management or size-selective regulations [12]. Although the capture of resilient species can maintain short-term yields, continued capture of ecologically valuable pelagic, demersal, crustacean, and elasmobranch species can lead to trophic decline, homogenisation, and disruption of ecosystem services [8, 11].
These findings emphasize the need for ecosystem-based management strategies, including spatial closures during critical life stages, adoption of bycatch reduction devices (BRDs), and modification of gear dimensions to minimize habitat overlap with non-target assemblages. Incorporating habitat-specific knowledge into gear design and effort allocation could enhance sustainability while maintaining economic viability in small-scale tropical bottom-trawl fisheries [30].
4.2 Community structure
The structure of the fish community in the study area was indicative of ecological imbalance commonly associated with intensive fishing pressure. Such assemblages have been characterized by low species diversity, uneven distribution of individuals, and disproportionate biomass dominance by one or two taxa, while others occur in marginal quantities [11, 24]. This condition reflects a state of ecological degradation that may compromise ecosystem functioning, including energy transfer, resilience, and regenerative capacity [9].
Compared to similar tropical systems, such as the open estuarine ecosystem along India's northwest coast [31], the mini-trawl catch in Lamongan exhibited greater ecological stress. The consistently low diversity and dominance of Stolephorus sp. suggest potential trophic shifts and intensifying exploitation, which could impair habitat structure and disrupt recovery processes [9, 32].
These findings reinforce the necessity of implementing ecosystem-based fisheries management that considers yield optimization, community structure, and ecosystem functioning. Recommended management strategies include the designation of protection zones, seasonal closures, gear-use restrictions, and the adoption of selective fishing technologies tailored to reduce ecological impacts while maintaining socio-economic viability [8, 9].
4.3 Variations in catch weight between species
The significant differences in mean catch biomass among species reflect the low selectivity of mini trawl gear, favouring the capture of small-bodied, schooling pelagic species. Stolephorus sp. contributed the highest biomass, consistent with its ecological traits—high abundance, tight schooling behaviour, and susceptibility to mass capture in tropical systems under intensive fishing pressure [23, 33]. These attributes raise concerns regarding long-term stock sustainability and trophic integrity.
In contrast, most demersal and pelagic fish, crustaceans, and molluscs exhibited no significant biomass variation and low average weights, indicating incidental capture of non-target species. Despite their limited contribution to total biomass, these species are functionally important within benthic ecosystems. Study [8] emphasised that non-selective gears such as trawls exert uniform pressure on slow-growing invertebrates and elasmobranchs, underscoring the need for targeted conservation measures.
The dominance of fast-growing, short-lived species, coupled with the scarcity of long-lived demersal predators, indicates potential ecosystem-level shifts consistent with the "fishing down the food web" phenomenon [28]. Technical modifications, including square mesh codends and bycatch reduction devices (BRDs), have been proposed to mitigate these impacts. Study [34] reported that such measures can reduce bycatch by up to 40% in tropical bottom trawl fisheries. Incorporating morphology- and size-based selectivity into gear design is critical to support sustainable exploitation and preserve functional community structure in coastal demersal systems.
4.4 Hierarchical Cluster Analysis (HCA) analysis
Hierarchical Cluster Analysis (HCA) based on morphological similarity identified three primary species groupings, reflecting shared structural traits (Figure 8). Cephalopods such as Sepia officinalis, Loligo sp., and Octopus sp. were clustered due to soft-bodied composition, bilateral symmetry, and similar body elongation ratios. Crustaceans, including Penaeus merguiensis and Charybdis anisodon, formed a separate cluster, highlighting their exoskeletal segmentation and locomotor adaptations. Meanwhile, demersal bony fishes such as Psettodes erumei and Lagocephalus spadiceus exhibited functional convergence, likely due to dorso-ventral flattening despite taxonomic divergence.
These morphological clusters carry significant implications for fishing gear selectivity. Species with similar body forms are more likely to pass through or be retained by similar mesh configurations, suggesting that body morphology can predict gear vulnerability. Studies [35, 36] demonstrated that integrating morphometric parameters—such as body length and girth—into net design improves species selectivity and reduces bycatch.
The broad linkage distances observed in the dendrogram reflect the low selectivity and wide functional range of mini trawl gear. These findings support the argument that morphology-informed fishing gear, including species-specific mesh panels or exclusion zones, is essential to mitigate incidental capture and preserve functionally diverse assemblages. Therefore, incorporating such morphological insights into gear development is a priority for sustainable trawl fisheries in tropical ecosystems [8, 30].
4.5 Integration of ANOVA and HCA results for fishing gear selectivity recommendations
The integration of ANOVA and HCA results highlights the low selectivity of mini trawl gear. ANOVA revealed significant differences in catch weight, with biomass dominated by Stolephorus sp. and Leiognathus splendens, while most demersal and crustacean species exhibited low and statistically similar catch weights. HCA further confirmed the gear's non-selectivity, as species with distinct morphologies and ecological roles—such as crustaceans (Charybdis anisodon, Penaeus merguiensis) and molluscs (Loligo sp.)—clustered alongside pelagic and demersal fishes, suggesting indiscriminate capture.
Based on these findings, three key recommendations emerge: (1) High-biomass species may be designated as target species; (2) Small, low-value species within shared morphological clusters could be excluded through gear modifications (e.g., square mesh codends, selective panels); and (3) Non-target taxa of conservation importance (e.g., crustaceans, molluscs, elasmobranchs) should be protected using bycatch reduction devices (BRDs) or excluder devices. Supporting evidence from study [34] demonstrates that square mesh panels and sorting grids can reduce non-target catch by up to 40%. Accordingly, mini trawl modifications in Lamongan should include increased mesh size, codend panel installation, time-area closures during spawning periods, and zoning adjustments in critical benthic habitats. These measures are expected to enhance economic efficiency, reduce bycatch, and mitigate ecological impacts on coastal fish communities.
4.6 General synthesis and ecosystem-based management implications
The findings of this study indicate that mini-trawl operations in Lamongan waters are strongly associated with the exploitation of small-bodied pelagic fishes such as Stolephorus sp. and Leiognathus splendens. At the same time, shrimps and cephalopods are consistently retained as bycatch. This catch structure is characterized by low gear selectivity, which simultaneously intercepts pelagic schooling fishes and demersal organisms. Consequently, ecological balance is diminished, recruitment overfishing is promoted, and economic inefficiencies are generated by capturing juveniles and non-target taxa.
Management recommendations should therefore be formulated by incorporating the morphological characteristics of the dominant bycatch species. The laterally compressed bodies of Leiognathus splendens and other ponyfishes suggest that use of square mesh with a condensate ≥25 mm is expected to enable the escape of undersized individuals while retaining larger target species [31]. Shrimps, characterized by rigid exoskeletons and elongated appendages, can be effectively released by applying rigid-grid BRDs or escape gaps positioned before the condend [32]. Cephalopods, including squids, possess flexible and elongated mantles that can be selectively excluded through separator panels providing vertical escape openings [33].
In addition to gear modification, temporal closures during the spawning seasons of the key taxa, such as shrimps, should be implemented to safeguard recruitment processes, while restrictions on trawl operations in shallow coastal habitats are expected to minimize the capture of juvenile pelagic fishes. These measures demonstrate that integrating species-specific morphological knowledge into technical gear design, supported by spatiotemporal management, provides a practical and implementable pathway to balance short-term fishing efficiency with long-term ecological sustainability. Such integration is consistent with ecosystem-based fisheries management (EBFM) principles, where ecological, biological, and socio-economic considerations are jointly incorporated into management frameworks [34, 35].
Mini-trawl fisheries in Lamongan waters were dominated by small pelagic fishes, particularly Stolephorus sp. and Leiognathus splendens, while shrimps, cephalopods, and demersal taxa constituted consistent bycatch. The catch structure was characterized by high richness but low diversity and evenness, reflecting ecological simplification and poor size selectivity that increase the risk of recruitment overfishing. Technical modifications, including square mesh codends, rigid-grid BRDs, and separator panels, together with seasonal closures and habitat restrictions, are recommended to mitigate these impacts. The integration of species-specific morphological knowledge into gear design is therefore essential for aligning catch efficiency with ecosystem-based sustainability.
The authors would like to thank all those involved in all stages of this research, from data collection, data processing, data analysis, and interpretation, to the writing of this publication.
[1] Akbari, N., Failler, P., Pan, H., Drakeford, B., Forse, A. (2023). The impact of fisheries on the economy: A systematic review on the application of general equilibrium and input–output methods. Sustainability, 15(7): 6089. https://doi.org/10.3390/su15076089
[2] FAO. (2024). The State of World Fisheries and Aquaculture - Blue Transformation in Action. Rome: FAO, Rome, Italy. https://doi.org/10.4060/cd0683en
[3] Mardhani, M., Syardiansah, S., Amilia, S., Aswadi, K. (2025). Assessing the contribution of marine fisheries to the indonesian economy. Journal of Coastal and Ocean Sciences, 6(1): 1-4. https://doi.org/10.31258/jocos.6.01.1-4
[4] Hilborn, R., Amoroso, R., Collie, J., Hiddink, J.G., Kaiser, M.J., Mazor, T., McConnaughey, R.A., Parma, A.M., Pitcher, C.P., Sciberras, M., Suuronen, P. (2023). Evaluating the sustainability and environmental impacts of trawling compared to other food production systems. ICES Journal of Marine Science, 80(6): 1567-1579. https://doi.org/10.1093/icesjms/fsad115
[5] Cömert, N., Deniz, T., Göktürk, D. (2025). Sustainable fisheries and non-target species management: A seasonal and depth-based study in the deep-sea fisheries of Antalya Bay. Sustainability, 17(11): 5040. https://doi.org/10.3390/su17115040
[6] Mytilineou, C., Herrmann, B., Smith, C.J., Mantopoulou-Palouka, D., et al. (2022). Impacts on biodiversity from codend and fisher selection in bottom trawl fishing. Frontiers in Marine Science, 9: 1021467. https://doi.org/10.3389/fmars.2022.1021467
[7] Kennelly, S.J., Broadhurst, M.K. (2021). A review of bycatch reduction in demersal fish trawls. Reviews in Fish Biology and Fisheries, 31(2): 289-318. https://doi.org/10.1007/s11160-021-09644-0
[8] Watkins, H.V., Munger, R.B., Côté, I.M. (2023). Effects of sea cucumber fishing on tropical seagrass productivity. Ecosphere, 14(5): e4543. https://doi.org/10.1002/ecs2.4543
[9] Tiano, J., De Borger, E., Paradis, S., Bradshaw, C., Morys, C., Pusceddu, A., Ennas, C., Soetaert, K., Puig, P., Masqué, P., Sciberras, M. (2024). Global meta-analysis of demersal fishing impacts on organic carbon and associated biogeochemistry. Fish and Fisheries, 25(6): 936-950. https://doi.org/10.1111/faf.12855
[10] Siddiqui, C., Rixen, T., Lahajnar, N., Lamont, T., Van Der Plas, A.K. (2024). Simulating potential impacts of bottom trawling on the biological carbon pump: A case study in the Benguela Upwelling System. Frontiers in Marine Science, 11: 1387121. https://doi.org/10.3389/fmars.2024.1387121
[11] Mazor, T., Pitcher, C.R., Rochester, W., Kaiser, M.J., et al. (2021). Trawl fishing impacts on the status of seabed fauna in diverse regions of the globe. Fish and Fisheries, 22(1): 72-86. https://doi.org/10.1111/faf.12506.
[12] Peixoto, U.I., Sporcic, M., Hobday, A.J., Bentes, B., Passarone, R., Fredou, F.L., Isaac, V.J. (2025). Ecological risk assessment of marine resources caught as bycatch in industrial bottom trawl shrimp fishery in the Amazon Continental Shelf. Frontiers in Marine Science, 12: 1490894. https://doi.org/10.3389/fmars.2025.1490894
[13] Ministry of Marine and Fisheries. (2015). Regulation of the Minister of Marine Affairs and Fisheries No. 2 of 2015 Concerning the Prohibition of the Use of Trawl Nets and Seine Nets in the Fisheries Management Area of the Republic of Indonesia. FAO. https://www.fao.org/faolex/results/details/es/c/LEX-FAOC197212.
[14] Susilowati, I., Suciati, I., Kusumawardhani, H.A., Sari, D.P.W., Puspita, S.D. (2024). The impact of the use of Sodo and Tiger Trawling Gear on communities in Tambakrejo Fishers' Village, Semarang. Jurnal Penelitian Ekonomi dan Bisnis, 9(1): 75-83. https://doi.org/10.33633/jpeb.v9i1.8145
[15] Nurhijayat, A., Adrianto, L., Booth, H.L., Boer, M., Zairion, Z., Salim, M.G., Yonvitner, Y., Tawang, F. (2025). Preliminary fisheries assessment and implications for sustainable blue economic development: A case study of Coastal Lamongan, East Java. BIO Web of Conferences, 176: 01003. https://doi.org/10.1051/bioconf/202517601003
[16] Carpenter, K.E., Niem, V.H. (1999). The living Marine Resources of the Western Central Pacific. Volume 3: Batoid Fishes, Chimaeras and Bony Fishes Part 1 (Elopidae to Linophrynidae). Food and Agriculture Organization of the United Nations. https://drive.google.com/drive/folders/1ApdCJkkTAAfo5H9E6ICv3kV1Ho4hsu_d.
[17] Carpenter, K.E., Niem, V.H. (2001). The Living Marine Resources of the Western Central Pacific. Volume 5: Bony Fishes Part 3 (Menidae to Pomacentridae). Food and Agriculture Organization of the United Nations. https://drive.google.com/drive/folders/1ApdCJkkTAAfo5H9E6ICv3kV1Ho4hsu_d.
[18] Carpenter, K.E., Niem, V.H. (1998). The Living Marine Resources of the Western Central Pacific. Volume 2: Cephalopods, Crustaceans, Holothurians and Sharks. Food and Agriculture Organization of the United Nations. https://drive.google.com/drive/folders/1ApdCJkkTAAfo5H9E6ICv3kV1Ho4hsu_d.
[19] Carpenter, K.E., Niem, V.H. (1999). The Living Marine Resources of the Western Central Pacific. Volume 4: Bony Fishes Part 2 (Mugilidae to Carangdae). Food and Agriculture Organization of the United Nations. https://drive.google.com/drive/folders/1ApdCJkkTAAfo5H9E6ICv3kV1Ho4hsu_d.
[20] Carpenter, K.E., Niem, V.H. (2001). The Living Marine Resources of the Western Central Pacific. Volume 6: Bony Fishes Part 4 (Labridae to Latimeriidae), Estuarine Crocodiles, Sea Turtles, Sea Snakes and Marine Mammals. Food and Agriculture Organization of the United Nations. https://drive.google.com/drive/folders/1ApdCJkkTAAfo5H9E6ICv3kV1Ho4hsu_d.
[21] Zar, J.H. (2014). Biostatistical Analysis, Fifth. Edinburgh: Pearson Education Limited.
[22] Montgomery, D.C. (2017). Design and Analysis of Experiments, Ninth. Arizona State: Wiley.
[23] Gilman, E., Hall, M., Booth, H., Gupta, T., Chaloupka, M., Fennell, H., Kaiser, M.J., Karnad, D., Milner-Gulland, E.J. (2022). A decision support tool for integrated fisheries bycatch management. Reviews in Fish Biology and Fisheries, 32(2): 441-472. https://doi.org/10.1007/s11160-021-09693-5
[24] Van Denderen, P.D., Plaza-Morlote, M., Vaz, S., Wijnhoven, S., et al. (2024). Complementarity and sensitivity of benthic state indicators to bottom-trawl fishing disturbance. Ecological Applications, 34(8): e3050. https://doi.org/10.1002/eap.3050
[25] Tiano, J.C., Depestele, J., Van Hoey, G., Fernandes, J., Van Rijswijk, P., Soetaert, K. (2022). Trawling effects on biogeochemical processes are mediated by fauna in high-energy biogenic-reef-inhabited coastal sediments. Biogeosciences, 19(10): 2583-2598. https://doi.org/10.5194/bg-19-2583-2022
[26] Yang, X., Luo, K., Fu, J., Kang, B., He, X., Yan, Y. (2025). Fish community resource utilization reveals benthic–pelagic trophic coupling along depth gradients in the Beibu Gulf, South China Sea. Biology, 14(2): 207. https://doi.org/10.3390/biology14020207
[27] Najmudeen, T.M., Sathiadhas, R. (2008). Economic impact of juvenile fishing in a tropical multi-gear multi-species fishery. Fisheries Research, 92(2-3): 322-332. https://doi.org/10.1016/j.fishres.2008.02.001
[28] Dineshbabu, A.P., Thomas, S., Josileen, J., Sarada, P.T., et al. (2022). Bycatch in Indian trawl fisheries and some suggestions for trawl bycatch mitigation. Current Science, 123(11): 1372-1380. https://doi.org/10.18520/cs/v123/i11/1372-1380
[29] Pauly, D., Christensen, V., Dalsgaard, J., Froese, R., Torres Jr, F. (1998). Fishing down marine food webs. Science, 279(5352): 860-863. https://doi.org/10.1126/science.279.5352.860
[30] Lucchetti, A., Melli, V., Brčić, J. (2023). Editorial: Innovations in fishing technology aimed at achieving sustainable fishing. Frontiers in Marine Science, 10: 1310318. https://doi.org/10.3389/fmars.2023.1310318
[31] Bhavan, S.G., Mujawar, S., Haragi, S., Rathod, J., Ingole, B., Mayekar, T., Lal, D.M., Murugesan, S.H., Bella, K., Appukuttannair, B., Sahadevan, P., Kutty, R., Kumar, P. (2023). A multi-metric fish index to measure the ecological quality of tropical predominantly open estuaries along the western coast of India. Environmental Monitoring and Assessment, 195(3): 421. https://doi.org/10.1007/s10661-023-11013-2
[32] Trindade-Santos, I., Moyes, F., Magurran, A.E. (2020). Global change in the functional diversity of marine fisheries exploitation over the past 65 years. Proceedings of the Royal Society B, 287(1933): 20200889. https://doi.org/10.1098/rspb.2020.0889
[33] Furuichi, S., Kamimura, Y., Suzuki, M., Yukami, R. (2022). Density-dependent attributes of schooling in small pelagic fishes. The Science of Nature, 109(5): 49. https://doi.org/10.1007/s00114-022-01819-4
[34] Jacques, N., Pettersen, H., Cerbule, K., Herrmann, B., Ingólfsson, Ó.A., Sistiaga, M., Larsen, R.B., Brinkhof, J., Grimaldo, E., Brčić, J., Lilleng, D. (2022). Bycatch reduction in the deep-water shrimp (Pandalus borealis) trawl fishery by increasing codend mesh openness. Canadian Journal of Fisheries and Aquatic Sciences, 79(2): 331-341. https://doi.org/10.1139/cjfas-2021-0045
[35] Vettiyattil, M., Herrmann, B., Bharathiamma, M. (2023). Square mesh codend improves size selectivity and catch pattern for Trichiurus lepturus in bottom trawl used along Northwest coast of India. Aquaculture and Fisheries, 8(5): 564-571. https://doi.org/10.1016/j.aaf.2021.12.015
[36] Santucci, R.G., Tosunoğlu, Z., Cilbiz, M., Charles, S.K., et al. (2024). Assessing codend mesh selectivity: Comparing diamond and square mesh codend in the Red Sea Shrimp Trawl Fishery of Saudi Arabia. Journal of Marine Science and Engineering, 12(10): 1848. https://doi.org/10.3390/jmse12101848