Association Study of IFNL3 and IFNLR1 Gene Polymorphisms with Systemic Lupus Erythematosus in Iraqi Women

Mohammed Amer Kamil*![]() | Hazima Mossa Al-Abassi
| Hazima Mossa Al-Abassi![]()
© 2025 The authors. This article is published by IIETA and is licensed under the CC BY 4.0 license (http://creativecommons.org/licenses/by/4.0/).
OPEN ACCESS
This study explored the association of IFNL3 (rs12979860) and IFNLR1 (rs4649203, rs72650431, rs72872528, rs59960858) polymorphisms with susceptibility to systemic lupus erythematosus (SLE) in Iraqi women. Ninety females were recruited: 30 untreated SLE patients, 30 treated patients, and 30 healthy controls. DNA was extracted from blood, and single-nucleotide polymorphisms (SNPs) were genotyped using PCR and sequencing. Analyses included odds ratios (ORs), 95% confidence intervals (CIs), Hardy–Weinberg equilibrium (HWE), and p-values. No SNP showed statistically significant association (p > 0.05), though some suggested protective or risk tendencies. For rs12979860, the CT genotype indicated a possible risk (OR = 1.18), while CC was protective (OR = 0.87). For rs4649203, GA and AA were protective (OR = 0.76 and 0.87), whereas the G allele suggested risk (OR = 1.41). rs72650431 showed AG as protective (OR = 0.72), while the G allele indicated risk (OR = 1.27). rs72872528 suggested GT as a risk (OR = 1.55) and the T allele pathogenic (OR = 2.63). For rs59960858, CA showed risk (OR = 1.17), while CC was protective (OR = 0.84). Notably, rs12979860, rs72650431, and rs72872528 deviated from HWE. Though not significant, these findings highlight possible roles for IFNL3 and IFNLR1 variants in SLE and warrant larger population-specific studies.
IFNL3, IFNLR1, SNP, polymorphism, SLP, Sanger sequencing, Hardy-Weinberg equilibrium, genotype
Systemic lupus erythematosus (SLE) is a chronic, multisystem autoimmune disorder characterized by dysregulated immune responses, autoantibody production, and systemic inflammation [1]. As a systemic immune disorder, SLE involves numerous organs when symptoms emerge [2]. Research indicates that disease development stems from the intricate interplay of genetic predisposition and environmental factors, which disrupts immune regulation and compromises immunological tolerance. SLE encompasses various autoimmune disorders with a range of clinical manifestations, including serositis, which involves the inflammation of serosal membranes that can lead to conditions like pericarditis, pleuritis, and peritonitis [3, 4]. Known for its relapsing nature, SLE symptoms can vary significantly in severity, from mild to life-threatening. The onset of SLE typically occurs during adolescence and early adulthood, predominantly affecting women aged 15 to 44 [5].
Interferons play a crucial role in SLE. These cytokines, produced in response to infections or inflammatory stimuli, are part of a broader family that possesses strong antiviral properties and is essential for regulating immune cell function. Type III consists of four subtypes and serves distinct immune functions. Notably, recent findings suggest that type III interferons, particularly interferon lambda, may also influence the pathogenesis of autoimmune and chronic inflammatory diseases, complicating the understanding of their roles. Crow [6] highlights SLE pathogenesis, emphasizing genetic, environmental, and immune factors, including interferon pathways, and B/T cell dysregulation, and identifies potential therapeutic targets for personalized treatment.
Interferons (IFNs) are essential cytokines that contribute significantly to effective antiviral defense. They are divided into three primary groups–types I, II, and III–according to their structural features, receptor engagement, and biological activities. While all IFNs are integral to antiviral defense, their specific roles differ significantly. Holicek et al. [7] reviewed type I interferon’s dual role in cancer, detailing its immune-stimulatory effects, tumor suppression, and context-dependent pro-tumor activities, informing therapeutic strategies. A landmark development in IFN research occurred in early 2003 with the discovery and initial characterization of the interferon lambda (IFN-λ) family. This group consists of three genes that encode closely related proteins designated as IFN-λ1, IFN-λ2, and IFN-λ3, which align with interleukin-29 (IL-29), IL-28A, and IL-28B, respectively. Al-Mashhadani et al. [8] reported elevated IL-1β, IFN-γ, and MMP-9 in Iraqi women with breast cancer, suggesting these mediators contribute to tumor progression and inflammatory responses. Consequently, type III IFNs (IFN-λs) exhibit expression patterns and biological actions similar to those of type I IFNs [9]. IFN-λ receptors are predominantly found on epithelial cells. This specificity underscores the prospective clinical importance of IFN-λ as an innovative antiviral treatment. Preliminary studies also indicate that IFN-λ might be advantageous in treating various medical conditions, including certain cancers [10]. Genetically, IFN-λ genes are located together on both murine chromosome 7 (in the 7A3 region) and human chromosome 19 (specifically in the 19q13.13 region) [11, 12].
Interferons drive aberrant immune activation, promoting the generation of autoantibodies and immune complexes that ultimately cause tissue injury. Research has linked SLE with dysregulation of both type I and type III interferons. Several studies, including those by study [13], reported elevated serum levels of IFNλ1 and IFNλ3 in SLE patients compared to healthy individuals. Additionally, the expression of IFNL1 transcripts has been found to increase in peripheral blood mononuclear cells (PBMCs) of SLE patients, while IFNL3 transcripts are elevated in activated CD4+ T cells [14, 15]. Significantly, higher serum concentrations of IFNλ are correlated with more severe disease manifestations and specific laboratory findings, such as elevated anti-double-stranded DNA (dsDNA) autoantibody titers and greater SLE Disease Activity Index values [16, 17]. Elevated IFNλ levels are also associated with various disease symptoms, such as arthritis, nephritis, serositis, and skin issues [13].
Volkova et al. [18] reported that a mechanistic QSP model incorporating IFN-I induction, signaling, and gene-signature readouts was used to compare the pharmacodynamics of anifrolumab, sifalimumab, daxdilimab, and litifilimab. This model provided testable predictions regarding the extent of IFN-signature suppression achieved by each therapeutic mechanism. Post-hoc proteomic and biomarker analyses from clinical trials further demonstrated that anifrolumab (anti-IFNAR1) effectively suppresses IFN-driven inflammatory proteins and pathways linked to disease activity, offering direct human evidence that IFN-I signaling causally drives pathogenic networks [19]. Complementary mechanistic dermatology studies and reviews highlight plasmacytoid dendritic cell (pDC) accumulation and IFN-I/MxA footprints in lesional skin, reinforcing the presence of tissue-localized IFN loops that parallel systemic disease [20].
Genetic studies have further elucidated the connection between genetic variations and the risk of SLE. Certain variants of IFNL3 and IFNL4 are linked to lupus nephritis in Taiwanese patients, while the rs4649203 single-nucleotide polymorphism in IFNLR1 is associated with an increased susceptibility to SLE among individuals from the Chinese Han population [21]. In a case study, serum IFNλ1 levels notably decreased during clinical remission following glucocorticoid and hydroxychloroquine treatment [22, 23]. Additionally, IFNλs have been shown to stimulate keratinocytes to express MHC class I molecules, potentially facilitating pathogenic CD8+ T cell responses [24].
This research sought to explore the potential association between genetic polymorphisms of the IFNL3 (rs12979860) gene and multiple polymorphisms in the IFNLR1 gene (rs4649203, rs72650431, rs72872528, and rs59960858) with systemic lupus erythematosus (SLE) incidence among Iraqi women.
2.1 Study population
The study involved 90 female participants, comprising both SLE patients and healthy controls, aged 20–40 years. Samples were obtained from Medical City (Consultant of Arthritis, Consultant of Dermatology, and the Hematology and Arthritis Lobby) at Baghdad Teaching Hospital between December 1, 2024, and March 1, 2025. Diagnosis was confirmed by specialists and the immunology unit of the aforementioned hospitals, following their established protocols and official records. The study sample consisted of 60 females with SLE, separated into two groups: 30 newly diagnosed, untreated patients (G2) and 30 patients receiving treatment (G3). The control group (G1) comprised 30 apparently healthy females. Individuals diagnosed with other illnesses, such as hypertension, cardiovascular disorders, and diabetes mellitus types I and II, or any other chronic autoimmune diseases and comorbid infections, were excluded from the study. Information collected for the samples included names, ages, residences, and occupations, which were documented through a structured questionnaire. Ethical authorization for this research was granted by the Scientific Research Committee of the Iraqi Ministry of Health.
2.2 Methodology
2.2.1 Blood sample collection
Each participant provided 5 ml of peripheral blood collected with disposable medical syringes after disinfecting the venipuncture site with 70% ethanol. Blood samples were aliquoted into 2.5 ml tubes for DNA extraction to investigate the genetic variations of IFNL3 and IFNLR1 using gene polymorphism analysis. The collected blood specimens were kept at 4℃ prior to use.
2.2.2 DNA extraction
An aliquot of 1 mL of blood sample was employed to isolate and purify genomic DNA from the participants with SLE and the control group using the EasyPure® Blood Genomic DNA kit, following the guidelines provided by the manufacturer. Agarose gel electrophoresis and nanodrop analysis were used to detect the integrity and concentration, respectively, of the isolated genomic DNA. The absorbance ratio (A260/A280) of the genomic DNA was calculated.
2.2.3 Polymerase chain reaction
In the current study, a PCR reaction was conducted for the patient and control group samples to amplify the IFNL3 and IFNLR1 genes. Genomic DNA was extracted from blood samples using the EasyPure® Blood Genomic DNA following the manufacturer’s instructions. Table 1 shows the sequences of the primers employed. Agarose gel electrophoresis was utilized to confirm the amplification. A 25 µL polymerase chain reaction mixture was prepared with approximately 10–100 ng of template DNA, 0.2 µM of each forward and reverse primer, 200 µM of each dNTP, 1X PCR buffer, 1.5–2.5 mM MgCl₂, and 1.25 U of Taq DNA polymerase. Nuclease-free water was added to make up the final volume. Amplification was performed with an initial denaturation at 95℃ for 3–5 min, followed by 30 cycles of denaturation at 95℃ for 30 s, annealing at 55℃ for 30 s, and extension at 72℃ for 30 s–1 min depending on the amplicon size. A final extension was carried out at 72℃ for 5–10 min, after which the reaction was held at 4℃ until further use. Amplified DNA fragments were stained using ethidium bromide before visualization using the Gel Documentation system.
Table 1. Sequences of the primers employed for the polymerase chain reaction
|
Primer Name |
Sequence (5’- 3’) |
Annealing Temp. (℃) |
Expected Product Size (bp) |
|
rs12979860-F2 |
GCGC TTAT CGCA TACG GCTA |
55 |
306 |
|
rs12979860-R2 |
TATG TCAG CGCC CACA ATTC |
||
|
rs4649203-F |
GTAAAACGACGGCCAGTCAAGGAGGTAGGTCAAAGTAAG |
1007 |
|
|
rs4649203-R |
CAGGAAACAGCTATGACCCCTCTCCTGTCTACTAAGATAA |
2.2.4 Statistical analysis
Data analysis was performed using the Statistical Package for the Social Sciences (SPSS, version 28). An independent t-test and one-way analysis of variance (ANOVA) with LSD post-hoc test were applied to determine p-values and significant differences among the study groups, while Pearson’s correlation coefficient was used to assess relationships within the data. All results were expressed as mean ± standard deviation (SD), with p < 0.05 considered statistically significant.
In the present study, the five SNPs were selected due to their roles in interferon signaling and autoimmune disease risk. IFNL3 rs12979860 is a functional haplotype marker influencing interferon-stimulated gene expression and linked to SLE susceptibility [25]. IFNLR1 rs4649203 shows genome-wide association with SLE in the Chinese Han population, while rs59960858 is a lead eQTL regulating IFNLR1 expression and colocalizing with psoriasis risk [26]. To ensure broader haplotype representation, rs72650431 and rs72872528 were chosen as tag variants within IFNLR1, following linkage disequilibrium–based selection strategies.
3.1 PCR-based detection of the IFNL3 gene
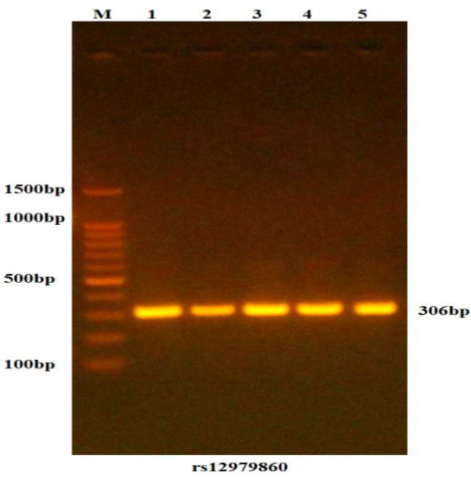
Agarose gel electrophoresis (Figure 1) and nanodrop analysis were used to detect the integrity and purity, respectively, of the genomic DNA isolated from the participants. The absorbance ratios of the DNA ranged from 1.8 to 2.0, indicating high purity. In the current study, a PCR reaction was conducted for the patient and control group samples to amplify the IFNL3 gene, and agarose gel electrophoresis was utilized to confirm this amplification for this region. Ethidium bromide staining of PCR amplified products revealed that all samples had a band and a fragment size of 306 bp. Figure 2 illustrates the agarose gel electrophoresis of the IFNL3 gene.

Figure 1. Gel electrophoresis of genomic DNA extracted from blood samples
Lanes 1-16 represent genomic DNA in 1% agarose gel at 70 volts for 60 min.
Figure 2. Amplification of the rs12979860-specific DNA region to detect the IFNL3 gene
DNA samples were separated on a 1.5% agarose gel pre-stained with ethidium bromide, and the gel picture revealed 306 bp PCR products; M: 100bp ladder marker; Lanes 1-5 resemble 306bp PCR products.
3.2 Association between rs12979860 polymorphism and SLE
Table 2 and Figure 3 present the genotype distribution and allele frequency of the SNP rs12979860. The distribution and allele frequency of the three genotypes, CC, CT, and TT, between SLE patients and control subjects did not differ significantly. The CT genotype functions as a risk factor (more susceptible to disease) because its odds ratio is greater than 1. However, the odds ratio was 0.87, and the CC genotype was significantly different; as a result, the CC genotype serves as a protective factor. In contrast, given that the odds ratio was 1, the TT genotype indicates that there is most likely no difference between the two groups. The odds ratio for the T allele was 1.08, while the odds ratio for the C allele was 0.91, indicating that the T allele could be associated with the susceptibility to the disease, while the C allele may act as a protective allele.

Table 2. Genotypic and allelic distribution of IFNL3 rs12979860 in study groups with Hardy-Weinberg equilibrium assessment
|
Group |
IFNL3 Polymorphism rs12979860 |
|||||
|
Genotype |
Allele |
|||||
|
CC |
CT |
TT |
C |
T |
||
|
Patients (60) |
No. |
34 |
18 |
8 |
86 |
34 |
|
% |
56.67% |
30% |
13.33% |
71.67% |
28.33% |
|
|
Control (30) |
No. |
18 |
8 |
4 |
44 |
16 |
|
% |
60% |
26.67% |
13.37% |
73.33% |
26.67% |
|
|
OR |
0.87 |
1.178 |
1.00 |
0.91 |
1.08 |
|
|
CI (95%) |
0.35-2.12 |
0.44-3.13 |
0.27-3.63 |
0.45-1.84 |
0.54-2.18 |
|
|
P value |
0.76 |
0.74 |
1.00 |
0.81 |
0.81 |
|
|
Hardy-Weinberg equilibrium for SNPs rs12979860 of the IFNL3 gene |
||||||
|
Group |
CC |
CT |
TT |
X2 |
P-value |
|
|
Control |
Observed |
18 |
8 |
4 |
3.037 C |
0.08 |
|
Expected |
16.13 |
11.73 |
2.13 |
|||
|
Patient |
Observed |
34 |
18 |
8 |
4.096 NC |
0.043* |
|
Expected |
30.82 |
24.37 |
4.82 |
|||
|
Total Observed |
|
52 |
26 |
12 |
- |
- |
Table 3. Comparative summary of key findings
|
Study Population |
Sample Size |
Key Finding |
Contrast with Iraqi study |
|
Current (Iraqi) |
60 patients, 30 controls |
No SLE association; HWE deviation in patients |
Reference |
|
Mexican [22] |
439 patients, 358 controls |
No SLE association; ↑ OASL in CC carriers |
Similar null association |
|
Taiwanese [22] |
164 SLE patients |
CC linked to lupus nephritis |
Ethnic divergence in end-organ risk |
|
Systemic Sclerosis [23] |
733 patients |
CC linked to pulmonary fibrosis (OR = 1.66) |
Highlights tissue-specific IFN effects |
|
HCV [24] |
191 patients |
CC predicts viral clearance (OR = 7.74) |
Opposite direction of effect |
These findings are supported by several related studies. For instance, it was reported that there was no association between rs12979860 and SLE susceptibility in Mexican individuals, but an increased OASL expression among CC carriers, which is consistent with the present study’s null overall association [27]. The Mexican study included a larger cohort of 439 patients compared with the 60 patients in the present study. Additionally, Beretta et al. [28] associated the CC genotype with pulmonary fibrosis in systemic sclerosis, which may explain the deviation from the Hardy-Weinberg equilibrium in the current patient group if fibrotic complications were present.
Several studies have demonstrated that the CC genotype favors viral clearance, which contrasts with its potential protective role in SLE, highlighting the pleiotropic effects of this variant [29, 30]. Furthermore, Hamdi et al. [31] suggested that CC enhances the expression of proinflammatory cytokines, potentially linking to the elevated OASL expression. The deviation from the Hardy-Weinberg equilibrium in patients may be attributed to factors such as selection bias (e.g., inclusion of patients with severe phenotypes), population stratification, or genetic drift. The control group being in HWE suggests no genotyping errors. The elevated CC frequency observed in both Iraqi groups (56-60%) contrasts with global patterns. While lower frequencies were reported in Mexican populations, Taiwanese studies identified associations with nephritis, underscoring the population-specific effects of this variant, as presented in Table 3 [27].
3.3 Polymerase chain reaction-detection of IFNLR1 gene
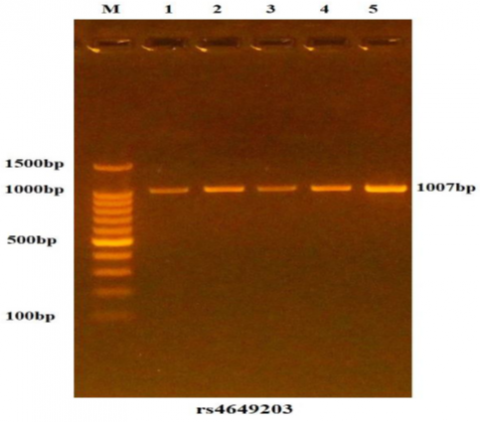
In this study, PCR was carried out on samples from both patient and control groups to amplify the IFNLR1 gene. The amplification was verified through agarose gel electrophoresis, as illustrated in Figure 4. The electrophoresis, performed with ethidium bromide staining, revealed a fragment size of 1007 bp, confirming successful amplification in all samples.
3.4 Association between rs4649203 polymorphism and SLE
As presented in Figure 5 and Table 4, the genotype distribution and allele frequency of SNP rs4649203 showed no significant differences among the three genotypes (GG, GA, and AA) between SLE patients and controls. The odds ratios for the GA and AA genotypes were 0.76 and 0.87, respectively, suggesting a potential protective effect of both genotypes. Notably, the GG genotype was absent in the control group. The odds ratio for the G allele was 1.41, whereas that for the A allele was 0.71, suggesting that the G allele may confer susceptibility to the disease, while the A allele may serve as a protective factor. Additionally, the outcomes in Table 4 show that the control group concurs with the HWE. There were no appreciable variations between the expected and observed genotypic frequencies. The HWE is compatible with both groups. Given that the total observed for both groups was 46, GA recorded a total of 38, and GG recorded a total of 6, indicating that the AA genotype may be regarded as a common genotype in the Iraqi female population.
Table 4. Genotypic and allelic distribution of IFNLR1 rs4649203 in study groups with Hardy-Weinberg equilibrium assessment
|
Group |
IFNLR1 Polymorphism rs4649203 |
|||||
|
Genotype |
Allele |
|||||
|
GG |
GA |
AA |
G |
A |
||
|
Patients (60) |
No. |
6 |
24 |
30 |
36 |
84 |
|
% |
10% |
40% |
50% |
30% |
70% |
|
|
Control (30) |
No. |
0 |
14 |
16 |
14 |
46 |
|
% |
0% |
46.67% |
53.33% |
23.33% |
76.67% |
|
|
OR |
- |
0.76 |
0.87 |
1.41 |
0.71 |
|
|
CI (95%) |
- |
0.31-1.84 |
0.36-2.11 |
0.68-2.87 |
0.34-1.45 |
|
|
P value |
- |
0.54 |
0.76 |
0.34 |
0.34 |
|
|
Hardy-Weinberg equilibrium for SNPs rs4649203 of the IFNLR1 gene |
||||||
|
Group |
GG |
GA |
AA |
X2 |
P value |
|
|
Control |
Observed |
0 |
14 |
16 |
2.78 C |
0.09 |
|
Expected |
1.63 |
10.73 |
17.63 |
|||
|
Patient |
Observed |
6 |
24 |
30 |
0.136 C |
0.712 |
|
Expected |
5.4 |
25.2 |
29.4 |
|||
|
Total Observed |
- |
6 |
38 |
46 |
- |
- |

As presented in Figure 6 and Table 5, the genotype distribution and allele frequency of SNP 72650431 showed no significant differences among the three genotypes (AA, AG, and GG) between SLE patients and controls. The odds ratio for the AG genotype was 0.72, indicating a possible protective effect, whereas the AA genotype showed no difference between the two groups, with an odds ratio of 1. Notably, the GG genotype was absent in the control group. The odds ratio for the G allele was 1.27, whereas that for the A allele was 0.78, suggesting that the G allele may be associated with increased disease susceptibility, while the A allele may function as a protective factor. The outcomes of the control groups agree with the HWE, as shown in Table 5. There were no appreciable variations between the expected and observed genotype frequencies. However, the HWE at p ≤ 0.007 was incompatible with the patient group. Other factors, such as selection for certain genotypes (i.e., the association signal itself) by the disease under study (SLE). Deviations from HWE in the control group, on the other hand, typically indicate population confounding or genotyping errors. Given that the total observed for both groups was 78, whereas AG recorded a total of 10 and GG recorded a total of 2, the results indicated that the AA genotype may be regarded as a common genotype in the Iraqi female population.

Table 5. Genotypic and allelic distribution of IFNLR1 72650431 in study groups with Hardy-Weinberg equilibrium assessment
|
Group |
IFNLR1 Polymorphism rs72650431 |
|||||
|
Genotype |
Allele |
|||||
|
AA |
AG |
GG |
A |
G |
||
|
Patients (60) |
No. |
52 |
6 |
2 |
110 |
10 |
|
% |
86.67% |
10% |
3.33% |
91.67% |
8.33% |
|
|
Control (30) |
No. |
26 |
4 |
0 |
56 |
4 |
|
% |
86.67% |
13.33% |
0% |
93.33% |
6.67% |
|
|
OR |
1.00 |
0.72 |
- |
0.78 |
1.27 |
|
|
CI (95%) |
0.27-3.63 |
0.18-2.78 |
- |
0.23-2.61 |
0.38-4.23 |
|
|
P value |
1.00 |
0.63 |
- |
0.69 |
0.69 |
|
|
Hardy-Weinberg equilibrium for SNPs rs72650431of the IFNLR1 gene |
||||||
|
Group |
AA |
AG |
GG |
X2 |
P-value |
|
|
Control |
Observed |
26 |
4 |
0 |
0.153 C |
0.69 |
|
Expected |
26.13 |
3.73 |
0.13 |
|||
|
Patient |
Observed |
52 |
6 |
2 |
7.16 NC |
0.007** |
|
Expected |
50.42 |
9.17 |
0.42 |
|||
|
Total Observed |
- |
78 |
10 |
2 |
- |
- |
3.6 Association between rs72872528 polymorphism and SLE
The genotypic distribution and allelic frequency for the SNP 72872528 are shown in Table 6 and Figure 7. The distribution of the three genotypes (GG, GT, and TT) and the frequency of each allele did not differ significantly between SLE patients and controls. The GG genotype had an odds ratio of 0.46. The GG genotype thus serves as a protective factor. But because the GT genotype has an odds ratio greater than 1 (1.55), it is a risk factor (more prone to disease). Conversely, the odds ratio for the TT genotype that was absent from the control group was (0). However, the odds ratio for the G allele was 0.37, and the odds ratio for the T allele was 2.63, suggesting that the T allele may be more likely to be linked to the illness. G, however, might serve as a protective allele. The Hardy-Weinberg equilibrium is supported by the results in Table 6 for the control groups. There were no appreciable variations between the expected and observed genotypic frequencies. The HWE was incompatible with the patient group. Other factors, such as selection for certain genotypes (i.e., the association signal itself) by the disease under study (SLE), could be the cause of this. Given that the total observed for both groups was 80, while GT recorded a total of 8 and TT recorded 2, the overall observed results imply that the GG genotype may be regarded as a common genotype in the Iraqi female population.

Table 6. Genotypic and allelic distribution of IFNLR1 72872528 in study groups with Hardy-Weinberg equilibrium assessment
|
Group |
IFNLR1 Polymorphism rs72872528 |
|||||
|
Genotype |
Allele |
|||||
|
GG |
GT |
TT |
G |
T |
||
|
Patients (60) |
No. |
52 |
6 |
2 |
110 |
10 |
|
% |
86.67% |
10% |
3.33% |
91.67% |
8.33% |
|
|
Control (30) |
No. |
28 |
2 |
0 |
58 |
2 |
|
% |
93.33% |
6.67% |
0% |
96.67% |
3.33% |
|
|
OR |
0.46 |
1.55 |
- |
0.37 |
2.63 |
|
|
CI (95%) |
0.09-2.33 |
0.29-8.21 |
- |
0.08-1.78 |
0.55-12.43 |
|
|
P value |
0.35 |
0.60 |
- |
0.22 |
0.22 |
|
|
Hardy-Weinberg equilibrium for SNPs rs72872528of the IFNLR1 gene |
||||||
|
Group |
GG |
GT |
TT |
X2 |
P-value |
|
|
Control |
Observed |
28 |
2 |
0 |
0.0357 C |
0.8 |
|
Expected |
28.03 |
1.93 |
0.03 |
|||
|
Patients |
Observed |
52 |
6 |
2 |
7.16 NC |
0.007** |
|
Expected |
50.42 |
9.17 |
0.42 |
|||
|
Total Observed |
- |
80 |
8 |
2 |
- |
- |
3.7 Association between rs59960858 polymorphism and SLE
The genotypic distribution and allelic frequency for SNP 59960858 are displayed in Figure 8 and Table 7. The distribution and allelic frequency of the three genotypes (CC, CA, and AA) between SLE patients and controls did not differ significantly. In the CC genotype, the odds ratio was 0.84. As a result, the CC genotype provides protection. On the other hand, the CA genotype acts as a risk factor (more susceptible to disease) because its odds ratio is greater than 1. Meanwhile, the control and patient groups' odds ratio was zero for the AA genotype. The A allele, on the other hand, may be more likely to be linked to the disease, as evidenced by the odds ratio of 1.14 for the A allele and 0.87 for the C allele. C, however, might serve as a protective allele. Additionally, Table 7 shows that the control group is consistent with the HWE. There were no significant variations between the expected and observed genotypic frequencies. The HWE is compatible with both groups. Because the total observed for both groups was 64, whereas CA recorded a total of 26 and GG recorded zero, the total observed results imply that the CC genotype may be regarded as a common genotype in the Iraqi female population.

Table 7. Genotypic and allelic distribution of IFNLR1 59960858 in study groups with Hardy–Weinberg equilibrium assessment
|
Group |
IFNLR1 Polymorphism rs59960858 |
||||||
|
Genotype |
Allele |
||||||
|
CC |
CA |
AA |
C |
A |
|||
|
Patients (60) |
No. |
42 |
18 |
0 |
102 |
18 |
|
|
% |
70% |
30% |
0% |
85% |
15% |
||
|
Control (30) |
No. |
22 |
8 |
0 |
52 |
8 |
|
|
% |
73.33% |
26.67% |
0% |
86.67% |
13.33% |
||
|
OR |
0.84 |
1.17 |
- |
0.87 |
1.14 |
||
|
CI (95%) |
0.31-2.26 |
0.44-3.13 |
- |
0.35-2.13 |
0.46-2.81 |
||
|
P value |
0.74 |
0.74 |
- |
0.76 |
0.76 |
||
|
Hardy-Weinberg equilibrium for SNPs rs59960858 of the IFNLR1 gene |
|||||||
|
Group |
CC |
CA |
AA |
X2 |
P-value |
||
|
Control |
Observed |
22 |
8 |
0 |
0.71 C |
0.4 |
|
|
Expected |
22.53 |
6.93 |
0.53 |
||||
|
Patient |
Observed |
42 |
18 |
0 |
1.87 C |
0.17 |
|
|
Expected |
43.35 |
15.3 |
1.35 |
||||
|
Total observed |
- |
64 |
26 |
0 |
- |
- |
|
Table 8. Summary of key SNP associations
|
SNP |
Genotype/Allele |
OR (95% CI) |
Role in SLE |
Consistent with HWE |
|
rs4649203 |
GA |
0.76 (0.31–1.84) |
Protective |
Yes (both) |
|
G allele |
1.41 (0.68–2.87) |
Risk |
||
|
rs72650431 |
AG |
0.72 (0.18–2.78) |
Protective |
No (patients) |
|
G allele |
1.27 (0.38–4.23) |
Risk |
||
|
rs72872528 |
GT |
1.55 (0.29–8.21) |
Risk |
No (patients) |
|
T allele |
2.63 (0.55–12.4) |
Risk |
||
|
rs59960858 |
CA |
1.17 (0.44–3.13) |
Risk |
Yes (both) |
|
A allele |
1.14 (0.46–2.81) |
Risk |
The results of this study focused on four SNPs (rs4649203, rs72650431, rs72872528, and rs59960858), analyzing their genotype and allele frequencies, odds ratios (ORs), confidence intervals (CIs), p-values, and HWE status (Table 8). For rs4649203, the GA and AA genotypes appeared potentially protective, whereas the G allele may represent a risk factor. In rs72650431, the AA genotype was predominant across both study groups, although a deviation from HWE was observed in the patient cohort. With respect to rs72872528, the GT genotype emerged as a possible risk factor, and the T allele may contribute to pathogenicity. Similarly, for rs59960858, the CA genotype was associated with increased risk, and the A allele was identified as a potential susceptibility factor. When placed in a broader biological context, evidence from recent studies underscores the role of IFNLR1 in antiviral immunity, particularly via activation of the JAK-STAT signaling pathway in epithelial and immune cells [32]. Given that dysregulated immune signaling and aberrant interferon responses are central to the pathogenesis of SLE, these findings suggest that genetic variation in IFNLR1 may have functional implications in disease susceptibility and progression. NCBI records confirm that IFNLR1 is a protein-coding gene expressed in immune-relevant tissues, supporting its potential role in autoimmune processes. Several studies have linked IFNLR1 variants to other immune-related conditions, including the association of rs4649203 with HCV treatment response [33], mutations in IFNLR1 with hearing loss [34], and reported links to pancreatic cancer. These associations reinforce the biological plausibility of the present findings. The findings are comparable with previous studies. In Iranian SLE cohorts, IL-1RN rs315952 shows a protective CT genotype and increased T-allele frequency in patients with hematologic manifestations, though not necessarily deviating from HWE [35]. Jordanian SLE patients exhibit strong associations with HLA-DRB1*0301, *1101, 1102, and DQB10601 alleles and risk haplotypes, which may reflect population stratification rather than technical error [36].
The functional effects of IFNL3/IFNL4 SNPs are well documented. rs12979860, linked to rs368234815, influences IFNL4 expression, affecting interferon-stimulated gene regulation and pro-inflammatory cytokine production, with the C allele enhancing viral clearance and immune activation [37, 38]. Additionally, rs59960858 modulates IFNLR1 expression, influencing autoimmune susceptibility [39].
The observed deviations from the Hardy-Weinberg equilibrium for rs72650431 and rs72872528 in patients may reflect population genetics factors such as consanguinity, which is particularly relevant within the Iraqi cohort. Deviations from the Hardy-Weinberg equilibrium (HWE) in patient groups suggest underlying biological or population processes. Such deviations may indicate selective pressures, genetic drift, inbreeding, or population stratification influencing allele distributions. In SLE, they may reflect disease-associated selection of certain genotypes, highlighting possible genetic susceptibility or resistance factors requiring further mechanistic and population-level investigations. The absence of significant associations may reflect the small sample size, population-specific genetic background, and deviations from the Hardy-Weinberg equilibrium. Additionally, disease heterogeneity, treatment effects, and stronger environmental or epigenetic influences could obscure genetic contributions, underscoring the need for larger, multi-ethnic studies to clarify IFNL3/IFNLR1 roles in SLE susceptibility.
This study found no statistically significant associations between IFNL3 (rs12979860) and IFNLR1 (rs4649203, rs72650431, rs72872528, rs59960858) polymorphisms and SLE susceptibility in Iraqi women, although some genotypes suggested modest risk or protective trends. Importantly, deviations from the Hardy-Weinberg equilibrium (HWE) observed in patient groups may reflect population-specific factors, such as consanguinity, genetic drift, or selection pressures associated with disease. These deviations highlight unique genetic dynamics within the Iraqi population and underscore the complexity of interpreting association studies in genetically structured cohorts. Future research should incorporate larger, multi-ethnic cohorts and functional analyses to validate the biological significance of these variants, while accounting for population stratification and environmental interactions in SLE pathogenesis.
The authors express their sincere thanks to the Department of Biology, College of Education for Pure Science (Ibn Al-Haitham), University of Baghdad, Iraq, for their assistance in the completion of this work.
[1] Kamil, M.A., Kadr, Z.H.M., Alabassi, H.M. (2022). Role of CXCL9-CXCR3 AXIS, ANA & DS-DNA ABS in Pathogenicity of SLE in Iraqi Patients. Pakistan Journal of Medical & Health Sciences, 16(4): 398-398.
[2] Ismael, A.K., Alabassi, H.M. (2024). The dynamic role of PD-1, vitamin D, RANKL, and sclerostin in Iraqi patients with systemic lupus erythematosus. Ibn AL-Haitham Journal For Pure and Applied Sciences, 37(1): 9-18. https://doi.org/10.30526/37.1.3367
[3] Accapezzato, D., Caccavale, R., Paroli, M.P., Gioia, C., Nguyen, B.L., Spadea, L., Paroli, M. (2023). Advances in the pathogenesis and treatment of systemic lupus erythematosus. International Journal of Molecular Sciences, 24(7): 6578. https://doi.org/10.3390/ijms24076578
[4] Lu, M.C., Hsu, C.W., Lo, H.C., Chang, H.H., Koo, M. (2022). Association of clinical manifestations of systemic lupus erythematosus and complementary therapy use in Taiwanese female patients: A cross-sectional study. Medicina, 58(7): 944. https://doi.org/10.3390/medicina58070944
[5] Al-Sarray, Z.A., Al-Rayahi, I.A., Al-Hafidh, A.H., Dwayyikh, A.T. (2020). Serum protein electrophoresis in Iraqi Systemic lupus erythematosus patient. Al-Nisour Journal for Medical Sciences, 2(1): 34-46. https://doi.org/10.70492/2664-0554.1048
[6] Crow, M.K. (2023). Pathogenesis of systemic lupus erythematosus: Risks, mechanisms and therapeutic targets. Annals of the Rheumatic Diseases, 82(8): 999-1014. https://doi.org/10.1136/ard-2022-223741
[7] Holicek, P., Guilbaud, E., Klapp, V., Truxova, I., Spisek, R., Galluzzi, L., Fucikova, J. (2024). Type I interferon and cancer. Immunological Reviews, 321(1): 115-127. https://doi.org/10.1111/imr.13272
[8] Al-Mashhadani, M.S.M., Al-Abassi, H.M., Mahood, W.S. (2022). The relationship between cellular mediators (IL-1β, IFN–γ, MMP-9) in Iraqi women with breast cancer. International Journal of Health Sciences, 6(S5): 1460-1474. https://doi.org/10.53730/ijhs.v6nS5.8982
[9] Alabassi, H.M., Al Hayani, D.A., Aljuamili, O.I., Ismael, A.K. (2023). The role of some immunological and hematological aspects in patients infected with COVID-19 in Al-Anbar province. Advances in Bioscience and Bioengineering, 11(2): 21-26. https://doi.org/10.11648/j.abb.20231102.11
[10] Liu, R.J., Yang, G.B. (2024). Molecular characteristics of rhesus macaque interferon-lambda receptor 1 (mmuIFNLR1): Sequence identity, distribution and alteration after simian-human immunodeficiency virus infection in the skin and buccal mucosa. Developmental & Comparative Immunology, 160: 105236. https://doi.org/10.1016/j.dci.2024.105236
[11] Al-Eqabi, H.F.K., Abd Al, S.A.A.H., Rahman, H.M. (2022). Dynamic role of IL-33, ST2 axis, SOD, leptin and nitric oxide in pathogenicity and disease progression in HBV chronic infection in Iraqi patients. Journal of Pharmaceutical Negative Result, 13(4): 258-263. https://doi.org/10.47750/pnr.2022.13.04.031
[12] Shahbazi, M., Amri Maleh, P., Bagherzadeh, M., Moulana, Z., et al. (2021). Linkage of lambda interferons in protection against severe COVID-19. Journal of Interferon & Cytokine Research, 41(4): 149-152. https://doi.org/10.1089/jir.2020.0187
[13] Adel, Y., Sadeq, Y. (2020). Impact of IL-34, IFN-α and IFN-λ1 on activity of systemic lupus erythematosus in Egyptian patients. Reumatologia, 58(4): 221-230. https://doi.org/10.5114/reum.2020.98434
[14] Oke, V., Gunnarsson, I., Dorschner, J., Eketjäll, S., Zickert, A., Niewold, T.B., Svenungsson, E. (2019). High levels of circulating interferons type I, II and III associate with distinct clinical features of active systemic lupus erythematosus. Arthritis Research & Therapy, 21: 107. https://doi.org/10.1186/s13075-019-1878-y
[15] Abdulridha, R.H., Saud, A.M., Alosami, M.H. (2022). Evaluation of interferon alpha (IFN-α) in women with systemic lupus erythematosus in Iraq. Iraqi Journal of Science, 63(10): 4225-4233. https://doi.org/10.24996/ijs.2022.63.10.9
[16] Yeo, A.L., Kandane-Rathnayake, R., Koelmeyer, R., Golder, V., et al. (2024). SMART-SLE: Serology monitoring and repeat testing in systemic lupus erythematosus—An analysis of anti-double-stranded DNA monitoring. Rheumatology, 63(2): 525-533. https://doi.org/10.1093/rheumatology/kead231
[17] Elsayed, S.A.R., Kamaly, H.M., Esmail, M.A. (2022). Co-positivity of anti-dsDNA, anti-nucleosome, and anti-smith autoantibodies as serological biomarkers for disease activity in systemic lupus erythematosus. Egyptian Rheumatology and Rehabilitation, 49(1): 8. https://doi.org/10.1186/s43166-021-00110-0
[18] Volkova, A., Sokolov, V., Tettamanti, F., Verma, M., Ugolkov, Y., Peskov, K., Tang, W., Kimko, H. (2025). An integrative mechanistic model of type 1 IFN-mediated inflammation in systemic lupus erythematosus. CPT: Pharmacometrics & Systems Pharmacology, 14(7): 1225-1235. https://doi.org/10.1002/psp4.70043
[19] Baker, T., Sharifian, H., Newcombe, P.J., Gavin, P.G., et al. (2024). Type I interferon blockade with anifrolumab in patients with systemic lupus erythematosus modulates key immunopathological pathways in a gene expression and proteomic analysis of two phase 3 trials. Annals of the Rheumatic Diseases, 83(8): 1018-1027. https://doi.org/10.1136/ard-2023-225445
[20] Hile, G.A., Werth, V.P. (2024). Understanding the role of type I interferons in cutaneous lupus and dermatomyositis: Toward better therapeutics. Arthritis Rheumatol, 77(1): 1-11. https://doi.org/10.1002/art.42983
[21] Li, Y., Cheng, H., Zuo, X.B., Sheng, Y.J., et al. (2013). Association analyses identifying two common susceptibility loci shared by psoriasis and systemic lupus erythematosus in the Chinese Han population. Journal of Medical Genetics, 50(12): 812-818.
[22] Bekaddour, N., Smith, N., Caspar, B., Grinberg, S., et al. (2024). The histamine analogue clobenpropit modulates IRF7 phosphorylation and interferon production by targeting CXCR4 in systemic lupus erythematosus models. Frontiers in Immunology, 15: 1490593. https://doi.org/10.3389/fimmu.2024.1490593
[23] Rasheed, R.A., Al-Abassi, H.M. (2021). TGF-Beta3 (gene expression) as a new biological marker for diagnosing the gastric and colorectal cancer. Biochemical and Cellular Archives, 21(1): 1539-1543.
[24] Mihaescu, G., Chifiriuc, M.C., Filip, R., Bleotu, C., et al. (2024). Role of interferons in the antiviral battle: From virus-host crosstalk to prophylactic and therapeutic potential in SARS-CoV-2 infection. Frontiers in Immunology, 14: 1273604. https://doi.org/10.3389/fimmu.2023.1273604
[25] Yilmaz, B., Çakmak Genç, G., Karakaş Çelik, S., Solak Tekin, N., Can, M., Dursun, A. (2022). Association between psoriasis disease and IFN-λ gene polymorphisms. Immunol Investigations, 51(6): 1772-1784.
[26] Wang, Y.F., Wei, W., Tangtanatakul, P., Zheng, L., et al. (2022). Identification of shared and Asian-specific loci for systemic lupus erythematosus and evidence for roles of type III interferon signaling and lysosomal function in the disease: A multi-ancestral genome-wide association study. Arthritis & Rheumatology, 74(5): 840-848. https://doi.org/10.1002/art.42021
[27] Juárez-Vicuña, Y., Pérez-Ramos, J., Adalid-Peralta, L., Sánchez, F., et al. (2021). Interferon Lambda 3/4 (IFNλ3/4) rs12979860 polymorphisms is not associated with susceptibility to systemic lupus erythematosus, although it regulates OASL expression in patients with SLE. Frontiers in Genetics, 12: 647487. https://doi.org/10.3389/fgene.2021.647487
[28] Beretta, L., Metwally, M., Thabet, K., Bayoumi, A., et al. (2020). IFNL3 genotype is associated with pulmonary fibrosis in patients with systemic sclerosis. Scientific Reports, 9(1): 14834.
[29] Ghodke-Puranik, Y., Olferiev, M., Crow, M.K. (2024). Systemic lupus erythematosus genetics: Insights into pathogenesis and implications for therapy. Nature Reviews Rheumatology, 20(10): 635-648. https://doi.org/10.1038/s41584-024-01152-2
[30] Lin, C.Y., Chen, J.Y., Lin, T.N., Jeng, W.J., Huang, C.H., Huang, C.W., Chang, S.W., Sheen,. I.S. (2011). IL28B SNP rs12979860 is a critical predictor for on-treatment and sustained virologic response in hepatitis C genotype-1 infection. PLoS One, 6(3): e18322. https://doi.org/10.1371/journal.pone.0018322
[31] Hamdi, E., Bekhit, A.A., Higazi, A., Ahmed, A.B.F., et al. (2023). Interferon-λ3 rs12979860 can regulate inflammatory cytokines production in pulmonary fibrosis. Saudi Pharmaceutical Journal, 31(11): 101816. https://doi.org/10.1016/j.jsps.2023.101816
[32] Novotny, L.A., Evans, J.G., Guo, H., Kappler, C.S., Meissner, E.G. (2024). Interferon lambda receptor-1 isoforms differentially influence gene expression and HBV replication in stem cell-derived hepatocytes. Antiviral Research, 221: 105779. https://doi.org/10.1016/j.antiviral.2023.105779
[33] Coto-Llerena, M., Lepore, M., Spagnuolo, J., Di Blasi, D., et al. (2021). Interferon lambda 4 can directly activate human CD19+ B cells and CD8+ T cells. Life Science Alliance, 4(1): e202000962. https://doi.org/10.26508/lsa.201900612
[34] Wang, W.Q., Qiu, S.W., Huang, S.S., Wang, G.J., Han, M.Y., Kang, D.Y., Yuan, Y.Y., Gao, X., Dai, P. (2022). Transcriptome analysis of the early stage IFNLR1-mutant zebrafish indicates the immune response to auditory dysfunction. Gene Expression Patterns, 43: 119229. https://doi.org/10.1016/j.gep.2021.119229
[35] Tahmasebi, Z., Akbarian, M., Mirkazemi, S., Shahlaee, A., Alizadeh, Z., Amirzargar, A.A., Jamshidi, A.R., Ghoroghi, S., Poursani, S., Nourijelyani, K., Mahmoudi, M. (2013). Interleukin-1 gene cluster and IL-1 receptor polymorphisms in Iranian patients with systemic lupus erythematosus. Rheumatology International, 33(10): 2591-2596. https://doi.org/10.1007/s00296-013-2784-2
[36] Khdair, S.I., Al-Bdour, R., Jarrar, W., Hammad, A., Al-Jayeh, A., Masa’deh, M., Adwan, M., Farah, R. (2022). Immunogenetic profiling of SLE and LN among Jordanian patients. Journal of Personalized Medicine, 12(12): 1955. https://doi.org/10.3390/jpm12121955
[37] Prokunina-Olsson, L., Muchmore, B., Tang, W., Pfeiffer, R.M., et al. (2013). A variant upstream of IFNL3 (IL28B) creating a new interferon gene IFNL4 is associated with impaired clearance of hepatitis C virus. Nature Genetics, 45(2): 164-171. https://doi.org/10.1038/ng.2521
[38] Balagopal, A., Thomas, D.L., Thio, C.L. (2010). IL28B and the control of hepatitis C virus infection. Gastroenterology, 139(6): 1865-1876. https://doi.org/10.1053/j.gastro.2010.10.004
[39] Rosenberg, B.R., Freije, C.A., Imanaka, N., Chen, S.T., et al. (2018). Genetic variation at IFNL4 influences extrahepatic interferon-stimulated gene expression in chronic HCV patients. The Journal of Infectious Diseases, 217(4): 650-655. https://pmc.ncbi.nlm.nih.gov/articles/PMC5853921/pdf/jix593.pdf.