Halimursyadah![]() | Syafruddin
| Syafruddin![]() | Siti Hafsah
| Siti Hafsah![]() | Rina Sriwati*
| Rina Sriwati*![]()
© 2025 The authors. This article is published by IIETA and is licensed under the CC BY 4.0 license (http://creativecommons.org/licenses/by/4.0/).
OPEN ACCESS
Patchouli (Pogostemon cablin Benth.) is a plant that yields essential oils with high economic value, contributing significantly to Indonesia’s economy, particularly in the perfume, cosmetics, and aromatherapy industries. However, patchouli oil production in Aceh has declined due to suboptimal cultivation practices. The application of PGPR (Plant Growth-Promoting Rhizobacteria) has the potential to enhance production by functioning as a biofertilizer, biostimulant, and bioprotectant. This study aimed to explore, isolate, characterize morphologically and physiologically, and apply PGPR to patchouli growth, identifying potential strains through 16S rRNA gene sequencing. The study began with the exploration of PGPR in the rhizosphere of patchouli plants across six regions in Aceh. Isolation and characterization were conducted at the Seed Science and Technology Laboratory, Faculty of Agriculture, Syiah Kuala University. PGPR was applied using a Randomized Block Design (RBD) with two factors: 15 rhizobacterial isolates and three Aceh patchouli varieties (Tapak Tuan, Lhokseumawe, and Sidikalang). This study identified 14 potential isolates capable of producing indole-3-acetic acid (IAA), fixing nitrogen, solubilizing phosphate, and generating Hydrogen Cyanide (HCN), siderophores, and 1-Aminocyclopropane-1-carboxylic acid (ACC) deaminase. Among them, isolate PG 9/2 C exhibited the highest efficacy and was identified as Delftia tsuruhatensis BB1455. This isolate enhanced plant height by up to 53.91% (Sidikalang), increased the number of leaves and branches by 89.88% and 85.56% (Tapak Tuan), improved root volume by 85.47% (Tapak Tuan), and boosted wet and dry biomass by 79.75% and 323.68% (Sidikalang), respectively, indicating optimal biomass accumulation.
Aceh patchouli, rhizobacteria, IAA, phosphate solubilization, nitrogen fixation
Patchouli (Pogostemon cablin Benth.) is a highly valuable aromatic plant, renowned for its essential oil, which is extensively utilized in the fragrance industry. Fluctuations in Aceh patchouli productivity over the past five years indicate challenges in cultivation management and limited land resources. In 2017, patchouli production reached 178 tons/ha, rising to 468, 1,219, 1,112, and 1,177 tons/ha in 2018, 2019, 2020, and 2021, respectively [1]. Given its significance as a key source of essential oil for Indonesia's export market, enhancing both the quality and yield of patchouli production has become a top priority [2]. However, most farmers have not implemented adequate cultivation interventions, leading to declining productivity. The practice of clearing new land or shifting cultivation is commonly adopted when the productivity of existing land declines, which can have adverse effects on environmental sustainability [2, 3]. To fulfill the increasing demand for patchouli oil, enhancing growth and yield through the application of Plant Growth-Promoting Rhizobacteria (PGPR) is essential.
PGPR are beneficial bacteria that inhabit plant roots and promote growth through various mechanisms, including nutrient solubilization, phytohormone production, and alleviation of environmental stress. Based on their location, PGPR can be classified into two categories: extracellular PGPR, which are found in the soil surrounding roots, root surfaces, and intercellular spaces, and intracellular PGPR, including nitrogen-fixing bacteria [4]. These bacteria enhance plant growth through direct mechanisms such as nutrient mobilization, nitrogen fixation, phosphate solubilization, and phytohormone production. Additionally, they indirectly modify microbial community composition and function in the rhizosphere, thereby promoting soil health [5, 6]. Rhizosphere engineering with PGPR can enhance soil structure, health, fertility, and function, thereby supporting plant growth in both normal and stressed conditions. PGPR also contributes to pathogen control by producing ACC deaminase, siderophores, antibiotics, lytic enzymes, and inducing systemic resistance, which in turn boosts plant tolerance to abiotic stress [5, 7].
Rhizobacteria can rapidly proliferate and use various substances as nutrient sources. Microbial diversity in rhizosphere soil is significantly higher compared to the roots, serving as the microbe-rich area. This happens because plants allocate a substantial amount of the carbon generated through photosynthesis to microbes around the roots, including mycorrhizal fungi and bacteria, to support the supply of nitrogen and phosphorus [8-11]. PGPR stimulates plants to produce various growth hormones, including auxins, cytokinins, and gibberellins, as well as volatile organic compounds. Additionally, PGPR produce growth regulators such as siderophores, which fix nitrogen, solubilize organic and inorganic phosphates, thereby enhancing nutrient availability and absorption by plants [4, 12]. Furthermore, PGPR increases plant tolerance to both biotic and abiotic stress, serving as an environmentally friendly alternative to chemical fertilizers, which can have negative impacts.
Several studies have identified various genera of rhizobacteria, including Azospirillum, Aeromonas, Azotobacter, Bacillus, Paenibacillus, Burkholderia, Enterobacter, Pantoea, Pseudomonas, and Rhizobium, which are capable of producing plant hormones [13]. Abscisic acid, cytokinins, gibberellins, IAA, and ethylene are essential phytohormones involved in plant-bacteria interactions. Research has identified Pseudomonas putida and Bacillus cereus as rhizobacteria that promote plant growth and offer resistance to Meloidogyne incognita, a root-knot nematode that affects patchouli. These bacteria also enhance overall patchouli growth and increase essential oil yield [14]. PGPR has also been proven to effectively enhance plant tolerance to abiotic stress and improve crop yields, serving as a promising biofertilizer [15].
Several studies have explored the potential of PGPR in promoting the growth of patchouli plants. Halimursyadah et al. [16] found that the rhizobacterial isolate PS 6/3 A from Purwosari Village, Nagan Raya Regency, helped increase the number of leaves and branches in patchouli plants. Further research by Halimursyadah et al. [16] identified multiple isolates, such as LB 4/2, LB 5/2 U, PT 7/2, and PT 8/2, which were effective in enhancing the growth of Aceh patchouli plants, particularly the Sidikalang variety, across various parameters including plant height, number of leaves and branches, leaf area, stem diameter, root length and volume, and wet biomass weight [17]. Previous studies were limited to the application of PGPR to patchouli with certain isolates and only focused on one plant variety and exploration of isolates without further analysis of the physiological and molecular characteristics that support their effectiveness as PGPR. This study addresses this gap by evaluating the effectiveness of 14 rhizobacterial isolates on three different patchouli varieties, namely Sidikalang, Lhokseumawe, and Tapaktuan, and conducting isolation, morphological and physiological characterization, and molecular identification using 16S rRNA on various PGPR isolates from six regions in Aceh. Although PGPR offer many benefits, numerous rhizobacteria associated with patchouli remain unexplored. As a result, this study was conducted to investigate, isolate, and characterize rhizobacteria from the rhizosphere of Aceh patchouli using physiological, agronomical, and molecular techniques.
2.1 Study area
Soil samples were collected using purposive sampling from the rhizosphere area of healthy plants at several smallholder patchouli plantations located in Aceh Province, as shown in Figure 1, which includes the following locations: Alue Abed, Calang (4°33'19.2"N95°45'43.4"E), Krueng Itam (4°01’02.9"N96°23’46.3"E), Purwosari (4°04’29.4"N 96°16’30.6"E), Lambada (5°30'45.6"N 95°35'40.5"E), Paya Tampu (4°46’23.5"N 96°21’58.8"E), Air Tenang (4°18’24.7"N 98°04’06.7"E, Tanah Terban (4°18’45.0"N 98°03’52.3"E), Bandar Mahligai Sekerak (4°16’28.4"N 98°01’23.0"E), Pulo Gelime (4°08’25.9"N 97°07’29.4"E).
Figure 1. Soil sampling map from the patchouli rhizosphere in several districts of Aceh Province
2.2 Isolation of rhizobacteria
In this study, the isolation method that was used for rhizobacteria was serial dilution. Initially, soil samples from several regions (Figure 1) were collected in 1 kg quantities from five different locations. Soil samples from each site were air-dried at room temperature (28-30℃) for 48 hours and then pooled [18]. After drying, the soil was sieved using a 9-mesh sieve shaker. One gram of the sieved soil was placed in a test tube, and 9 ml of distilled water was added. The mixture was vortexed at 100 rpm to create a uniform solution, which was labelled as the first dilution (10⁻¹). A 1 mL sample of the 10⁻¹ dilution was transferred to a new test tube with 9 ml of dilution solution, creating the second dilution (10⁻²). This process was repeated until the final dilution of 10⁻⁹. Nutrient Agar (NA) was prepared in Petri dishes, allowed to solidify, and used to isolate rhizobacteria. A 1 mL sample from dilutions 10⁻⁴ to 10⁻⁹ was plated in duplicate on NA agar plates and incubated at 28℃ for 48 hours. After incubation, bacterial identification was carried out, and pure rhizobacterial cultures were obtained.
2.3 Physiological identification
2.3.1 IAA production test
The Glickmann and Dessaux approach was used to assess the rhizobacterial isolates' capacity to generate IAA. The isolates were cultured on Sucrose Peptone Agar (SPA) medium for a whole day [19]. Each media received 0.5 g/l of the amino acid tryptophan to promote auxin production, which was then centrifuged for 10 minutes at 10,000 rpm. After being removed from the bacterial pellet, the supernatant (centrifugation liquid) was filtered through 0.2 μm Millipore filter paper, and its IAA level was measured. Salkowski's reagent was used to determine the amount of IAA present in the bacterial culture filtrate. A 2 ml test tube or an Eppendorf tube was filled with 1 ml of the bacterial culture filtrate and 1 ml of Salkowski's reagent. The absorbance was then measured using a spectrophotometer calibrated at a wavelength of 550 nm after 30 minutes of incubation in the dark at 26℃. To determine the amount of IAA in the bacterial culture filtrate, a standard curve based on the absorbance values of pure IAA solutions with concentrations of 0, 6.25, 12.5, 25, 50, 75, 100, 150, and 200 µg/ml was employed [20].
2.3.2 Phosphate solubilizing ability test
Phosphate source in an agar-based medium to examine the rhizobacteria's capacity to solubilize phosphate. In 1000 milliliters of distilled water, 15 grams of agar, 10 grams of glucose, 0.2 grams of KCl, 0.2 grams of NaCl, 5 grams of Ca₃(PO₄)₂, 0.5 grams of yeast extract, 2.5 grams of MnSO₄, 2.5 grams of FeSO₄, and 0.1 grams of MgSO₄ were dissolved to create the medium, which was then autoclaved to ensure sterilization. Using a cork borer, three wells were created in the test medium before it was transferred into Petri dishes. 0.2 ml of the rhizobacterial suspension under test was added to each well, and the wells were then incubated for seven days at 28℃. According to study [21], the development of a clean zone surrounding the well holding the bacterial suspension demonstrated the rhizobacteria's capacity to solubilize phosphate. The Phosphate Solubilizing Index (PSI) was computed using the clear zone that developed around each isolate. Following seven days of incubation, measurements were made of the colony's and the clear zone's diameters [22]. The following formula was used to determine the PSI:
$P S I=\frac{A+B}{B}$ (1)
where, A=Diameter clear zone (cm), B=Colony diameter (cm), PSI=Phosphate Solubilizing Index.
2.3.3 Nitrogen fixation ability test
A medium including MgSO₄·7H₂O (25 g), FeSO₄·7H₂O (0.01 g), NaMoO₄·2H₂O (0.01 g), MnSO₄·5H₂O (0.01 g), CaCl₂ (0.1 g), K₂HPO₄ in NaCl buffer (5 mL), and Agar (17–20 g) was used to examine the nitrogen fixing ability of rhizobacteria isolates. Pure isolates that were 48 hours old were cultivated on the medium, incubated for four days at room temperature (28℃), and the growth of the bacteria was monitored. The growth of the isolate in nitrogen-free media, as demonstrated by the shift in the liquid medium's turbidity, served as a signal for isolates that could fix nitrogen [20].
2.3.4 Hydrogen Cyanide (HCN) production test
A technique proposed by Bakker and Schipper [23] was used to qualitatively examine the formation of hydrogen cyanide (HCN) by rhizobacteria. 4.4 g of glycine, 2 g of picric acid, 8 g of sodium carbonate, 15 g of agar, 30 g of TSB, 1000 mL of distilled water, and pieces of sterile filter paper were used to prepare the testing medium. After dissolving glycine, TSB, and agar in 1000 milliliters of sterile distilled water and autoclaving the solution, it was transferred into Petri dishes. Two grams of picric acid and eight grams of sodium carbonate were dissolved in 200 milliliters of sterile distilled water to prepare the HCN detection solution. Sterile filter paper pieces were soaked in this solution before being placed in the Petri dish lid, and bacteria were then streaked onto the glycine medium. The filter papers were then placed in the center of the Petri dish lid. The plates were incubated at 28℃ for seven days. During incubation, HCN production by rhizobacteria was qualitatively observed. Isolates that did not cause any color change in the filter paper indicated no HCN production. HCN-producing isolates induced the following color changes in the filter paper: from yellow to light brown (+HCN low), light brown to dark brown (++HCN moderate), dark brown to brick red (+++HCN high), and dark red to black (++++HCN very high).
2.3.5 Siderophore production test
The production of siderophores by rhizobacterial isolates was tested by culturing the bacteria in Chrome Azurol S (CAS) medium, a standard method for detecting and screening siderophore producers. The test was carried out for 24 hours at room temperature. The medium was prepared with the following composition per liter: 20 g sucrose, 2 g L-asparagine, 1 g K₂HPO₄, and 0.5 g MgSO₄. After centrifuging rhizobacterial suspensions for 30 minutes at 11,000 rpm, the supernatant was filtered through a 0.2 µm nitrocellulose membrane. A control sample of supernatant without FeCl₃ was compared to 3 mL of the supernatant after 1 mL of 0.01 M FeCl₃ was added to evaluate siderophore formation. A spectrophotometer set at 410 nm was used to measure the synthesis of siderophores [24].
2.3.6 ACC-Deaminase production test
Following the protocol of Glick [25], ACC-Deaminase activity was assessed using Dworkin-Foster (DF) minimal salt medium supplied with the substrate 1-aminocyclopropane-1-carboxylate (ACC) or ammonium sulfate as the nitrogen source. Four grams of KH₂PO₄, six grams of Na₂HPO₄, 0.2 grams of MgSO₄·7H₂O, one milligram of FeSO₄·7H₂O, ten μg H₃BO₃, ten μg MnSO₄, seventy μg ZnSO₄, fifty μg CuSO₄, ten μg MoO₃, two grams of glucose, two grams of gluconic acid, two grams of citric acid, and twelve grams of agar (for solid media) were all dissolved in 1000 milliliters of distilled water. To the DF medium, 2 g of ammonium sulfate was added. All parts were then autoclaved for 30 minutes at 121℃ and 0.1 MPa to disinfect them. This was followed by inoculation with 1 mL of bacterial suspension (1 × 10⁹ cells/mL) onto both DF and DF + ammonium sulfate media. Growth of the isolates was observed after 48 hours, following the method described by the study [26]. Isolates that grew on DF + ammonium sulfate showed the presence of ACC-Deaminase activity. The DF medium without ammonium sulfate was used as a control to determine when the isolates were nitrogen-fixing bacteria capable of obtaining nitrogen by fixing N₂ from the air.
2.3.7 Analysis of soil nutrient content
In this study, soil was analyzed at the Agricultural Instrument Standardization Application Center's Testing Service Laboratory in Aceh Province before and after the application of rhizobacteria. Mon Alue in Aceh Besar's Indrapuri District provided the soil utilized as the planting medium. K-available (ppm), K-exchangeable, Na-exchangeable, Ca-exchangeable, Mg-exchangeable, C-Organic (%), N-Total (%), C/N ratio, pH H₂O, and Cation Exchange Capacity (CEC) (meq/100g) were among the soil metrics [27].
The organic carbon and total nitrogen content of the soil are measured to perform the carbon-to-nitrogen (C/N) ratio analysis. 0.5 g of air-dried soil sample is reacted with 1 N potassium dichromate (K₂Cr₂O₇) and concentrated sulfuric acid (H₂SO₄), shaken, and left to stand before being titrated with 0.5 N ferrous sulfate (FeSO₄) using ferroin indicator until the color changes from blue-green to red-brown. This is the Walkley and Black [28] method for determining organic carbon. Meanwhile, total nitrogen analysis follows the Kjeldahl method (1883) where 0.5 g of soil sample is digested with concentrated H₂SO₄ and a catalyst, then distilled with 40% NaOH to convert ammonium (NH₄⁺) into ammonia gas (NH₃), which is then trapped in 4% boric acid solution and titrated with 0.1 N H₂SO₄ or HCl using bromocresol green and methyl red indicator [29]. The C/N ratio is determined by dividing the organic carbon content by the total nitrogen content, serving as an indicator of the organic matter decomposition rate in the soil.
Cation exchange capacity (CEC) was analyzed using the ammonium acetate method at pH 7.0 [30]. A 5 g air-dried soil sample was placed in a centrifuge tube and mixed with 25 mL of 1 N ammonium acetate (CH₃COONH₄) solution at pH 7.0. The mixture was shaken for 30 minutes to exchange the cations adsorbed on soil colloids with ammonium ions (NH₄⁺). The solution was then separated from the soil by filtration or centrifugation, and the filtrate was analyzed to determine the concentrations of exchangeable cations such as calcium (Ca²⁺), magnesium (Mg²⁺), potassium (K⁺), and sodium (Na⁺). Inductively Coupled Plasma-Optical Emission Spectrometry (ICP-OES) or Atomic Absorption Spectroscopy (AAS) was used to quantify these cations. The total amount of exchangeable cations in cmol(+) kg⁻¹ soil was used to calculate CEC [31].
2.4 Agronomic characterization
2.4.1 Preparation of growing media and plant material
The initial step included preparing the planting medium by mixing topsoil and compost at a 1:1 ratio. For two weeks, this soil mixture was sterilized using 20 g m-3 of the soil sterilant Basamid G. A pH meter was used to measure the medium's pH after sterilization; 6-7 is the ideal pH range. Subsequently, planting materials were prepared in the form of stem cuttings with 5-6 leaves from each variety. The varieties used in this study were Tapak Tuan, Lhokseumawe, and Sidikalang. The stem cuttings are initially propagated for 21 to 30 days using the sungkup (humidity chamber) method. This method ensured uniform and healthy growth of the patchouli seedlings.
2.4.2 Preparation of rhizobacteria suspension
Fifteen rhizobacteria isolates from various regions that showed superior physiological performance in preliminary tests, were selected to investigate their effects on patchouli plant growth. Without coming into contact with the growing medium, these isolates were cultivated and suspended in 100 milliliters of sterile distilled water. Using a spectrophotometer, the population density of the suspension was calculated to achieve a concentration of 109 CFU mL-1, corresponding to an absorbance of OD600 = 0.192 [32]. Subsequently, the bacteria suspension was transferred to culture bottles for application.
2.4.3 Planting and application of rhizobacterial isolates
After four weeks of growth, the patchouli seedlings were ready for transplantation into polybags measuring 40 × 25 cm, containing a 2:1 mixture of soil and compost with a total volume of 10 kg. The soil used for planting was obtained from the patchouli rhizosphere region. Following transplantation, a suspension of indigenous rhizobacteria was applied to the seedlings. The inoculation was carried out once a week after transplantation, with 100 mL of the bacterial suspension applied around the root zone of each plant. Other maintenance activities included regular watering and weed, pest, and diseases control to ensure healthy growth.
2.5 Molecular identification and phylogenetic tree construction
One popular technique in molecular biology is the polymerase chain reaction (PCR). PCR is widely used in molecular analysis, especially for amplifying specific regions (amplicons) and identifying specific genes within a sample. It is based on the ability of DNA polymerase to synthesize a new DNA strand from a given template. In molecular biology, real-time PCR, also known as quantitative real-time PCR (qPCR), is becoming increasingly popular. qPCR can be used to detect the presence of a specific gene, compare expression levels across different samples, and quantify gene abundance. In this study, DNA extraction from rhizobacteria was performed using the DNeasy Blood and Tissue Kit (https://ptgenetika.com/Lab. Genetika, Banten). After being moved to an Eppendorf tube with 500 µL of buffer solution, the genomic DNA was crushed with a grinder. After centrifuging the sample for ten minutes at 13,000 rpm, the supernatant was carefully poured into a fresh Eppendorf tube. Subsequently, 450 µL of a 24:25:1 Phenol: Chloroform: Isoamyl alcohol (PCI) solution was added, and the mixture was shaken for five minutes. After centrifuging the mixture for ten minutes at 13,000 rpm, the supernatant was carefully transferred into a new Eppendorf tube. Finally, to precipitate the DNA, 500 µL of cold 100% ethanol was added [27].
2.6 Observation parameters
The study's characteristics included the rhizobacterial isolates' capacity to fix nitrogen, solubilize phosphate, generate IAA, and create the enzyme ACC deaminase. At 12 weeks following application (WAA), measurements were also made of the plant's height (cm), number of leaves, number of branches, root volume, wet weight (g), and dry weight (g).
At 30, 60, and 90 days after application (DAA), established techniques were used to measure the plant growth characteristics. While the number of leaves and branches was counted based on completely formed organs, the height of the plant was measured from the base of the stem to the growing point. At 90 DAT, the leaf area was examined using ImageJ software. Following harvesting and drying, fresh and dry biomass were weighed using a digital balance. A ruler was used to measure the length of the roots, and a graduated cylinder's water displacement method was used to calculate the root volume.
2.7 Data analysis
This study employed a randomized block design (RBD) with a factorial arrangement. The first factor was the type of indigenous rhizobacteria (R) sourced from a prior study. The identification results indicated that these indigenous rhizobacteria functioned as biofertilizers and biostimulants, as demonstrated by their IAA production and phosphate solubilization abilities. This factor included 15 levels: R0 =no rhizobacteria (control), and R1 to R14. The second factor was the variety (V), which had three levels: V1 =Lhokseumawe variety, V2 =Tapak Tuan variety, and V3 =Sidikalang variety. As a result, there were 45 treatment combinations, replicated three times, leading to 135 experimental units, each consisting of two plants, totalling 270 samples. Data were analyzed using Analysis of Variance (ANOVA), and when the F-test showed significant effects (α =5%), further analysis was performed using Duncan's New Multiple Range Test (DNMRT) at the 5% significance level.
3.1 Physiological characterization of rhizobacteria as PGPR
The potential of rhizobacteria isolates as PGPR was shown by their ability to produce phytohormones such as IAA, solubilize phosphate, fix nitrogen, and produce the enzyme ACC deaminase. The results of the rhizobacteria isolates PGPR activity are presented in Table 1.
Table 1. Physiological characterization of indigenous rhizobacterial isolates from the patchouli rhizosphere in several districts of Aceh Province
|
Isolate Code |
IAA (ppm) (y=0,018x-0,001) |
Phosphate Solubilization Index |
Siderophore Production (After FeCl Addition) |
Nitrogen Fixation |
HCN Production |
ACC-Deaminase Production |
|
AT 7/1 C |
26.43 ± 1.77fg |
4.26 ± 0.31l |
0.47 ± 0.00kl |
++ |
++ |
++ |
|
AT 7/1 K |
23.17 ± 1.09cde |
3.46 ± 0.38fghij |
0.48 ± 0.01kl |
+ |
+ |
+ |
|
BMS 8/3 P |
22.04 ± 2.97bcd |
3.30 ± 0.24defgh |
0.49 ± 0.01l |
++ |
+++ |
++ |
|
BMS 8/5 H |
24.13 ± 1.36cdef |
2.10 ± 0.10b |
0.52 ± 0.01m |
++ |
++ |
++ |
|
CL 4/1 |
44.17 ± 0.31l |
3.27 ± 0.15defgh |
0.28 ± 0.02g |
- |
+ |
+ |
|
CL 8/1 |
24.31 ± 0.04cdef |
2.73 ± 0.03c |
0.24 ± 0.00de |
- |
- |
+ |
|
KI 8/1 |
70.63 ± 0.25n |
3.27 ± 0.15defgh |
0.38 ± 0.01j |
++ |
++ |
+ |
|
KI 8/3 |
42.28 ± 0.29kl |
3.76 ± 0.34ijk |
0.37 ± 0.02j |
- |
- |
+ |
|
KI 8/4 |
39.61 ± 0.68jk |
3.35 ± 0.21defghi |
0.22 ± 0.03cd |
- |
++ |
++ |
|
LB 4/1 C |
20.00 ± 2.79b |
3.60 ± 0.02ghij |
0.36 ± 0.02ij |
- |
- |
++ |
|
LB 4/2 |
28.83 ± 3.23g |
4.05 ± 0.22kl |
0.25 ± 0.01def |
+ |
- |
++ |
|
LB 5/1 K |
15.85 ± 2.97a |
3.02 ± 0.17cdef |
0.34 ± 0.03h |
- |
+ |
++ |
|
PG 7/2 |
24.65 ± 0.22cdef |
3.01 ± 0.13cdef |
0.26 ± 0.01efg |
+ |
- |
+ |
|
PG 7/3 CK |
15.00 ± 0.31a |
3.67 ± 0.17hijk |
0.49 ± 0.03l |
- |
- |
+ |
|
PG 8/1 |
14.20 ± 0.98a |
3.19 ± 0.13defg |
0.19 ± 0.02bc |
+ |
- |
++ |
|
PG 9/2 C |
14.54 ± 1.65a |
2.72 ± 0.10c |
0.45 ± 0.02k |
- |
- |
++ |
|
PS 4/2 |
42.87 ± 0.26 l |
3.33 ± 0.23defghi |
0.27 ± 0.02efg |
+ |
++ |
+ |
|
PS 4/5 |
52.44 ± 0.40m |
0.00 ± 0.00a |
0.18 ± 0.01b |
+ |
++ |
+ |
|
PS 5/6 C |
82.02 ± 1.78o |
2.99 ± 0.17cde |
0.17 ± 0.01b |
- |
+++ |
- |
|
PS 5/6 PK |
36.37 ± 1.24hi |
2.89 ± 0.23cd |
0.17 ± 0.02b |
- |
+ |
+ |
|
PS 6/1 |
33.87 ± 1.01h |
3.13 ± 0.22cdef |
0.29 ± 0.01g |
++ |
++ |
+ |
|
PS 6/3 A |
36.69 ± 0.69hi |
2.69 ± 0.40c |
0.32 ± 0.02h |
+ |
++ |
++ |
|
PS 6/5 |
37.44 ± 0.64hi |
3.81 ± 0.23jk |
0.24 ± 0.01de |
++ |
++ |
+ |
|
PT 7/2 |
21.56 ± 2.37bc |
4.78 ± 0.40m |
0.73 ± 0.02n |
++ |
+ |
- |
|
PT 8/1 |
27.20 ± 2.40fg |
4.24 ± 0.31l |
0.28 ± 0.01fg |
++ |
- |
++ |
|
PT 8/2 |
15.24 ± 2.25a |
3.41 ± 0.28efghij |
0.28 ± 0.02g |
++ |
+ |
++ |
|
TT 8/3 CR |
24.72 ± 2.56def |
3.03 ± 0.22cdef |
0.14 ± 0.01a |
++ |
+++ |
++ |
|
TT 8/4 P |
26.09 ± 1.77efg |
4.32 ± 0.40l |
0.24 ± 0.02de |
+ |
+ |
++ |
Note: The first two letters in the isolate code indicate the origin of the isolate, the number represents the dilution factor used, and the last letter of the code denotes the colony color.
Table 1 shows that all rhizobacteria isolates from various patchouli rhizospheres are capable of producing IAA. The ability of these isolates to act as potential PGPR was shown by their production of IAA, a phytohormone capable of stimulating plant growth. Based on the results, isolate PS 5/6 C showed the highest IAA production, with a value of 82.02 ± 1.78 ppm, followed by KI 8/1 with 70.63 ± 0.25 ppm. Other isolates showing high potential include CL 4/1 and PS 4/5 with 44.17 ± 0.31 ppm and 52.44 ± 0.40 ppm, respectively. Most isolates showed varying IAA levels, with PG 9/2 C, PG 7/3 CK, and LB 5/1 K, producing IAA levels below 30 ppm, indicating a lower production capacity. These results suggested that isolates with high production could serve as growth-promoting agents, potentially enhancing plant growth and productivity by stimulating root development and improving nutrient absorption.
Based on the data presented regarding the phosphate solubilization index of various rhizobacterial isolates, isolate PT 7/2 showed the highest phosphate solubilization capacity with an index of 4.78 ± 0.40, followed by AT 7/1 C, TT 8/4 P, and PT 8/1 with values of 4.26 ± 0.31, 4.32 ± 0.40, and 4.24 ± 0.31, respectively. Most isolates had phosphate solubilization indices above 3.00, showing their ability to improve phosphate availability to plants. However, PS 4/5 exhibited no phosphate solubilizing activity, with a value of 0.00 ± 0.00. This indicates that not all isolates had the same capacity to solubilize phosphate. Therefore, isolates with a high phosphate solubilization index could be used as growth-promoting agents by enhancing the availability of essential nutrients including phosphate. Isolates PT 7/2 and BMS 8/3 P showed high siderophore production, with values of 0.73 and 0.52, respectively. Both isolates could contribute to iron binding, thereby enhancing nutrient availability to plants. While TT 8/3 CR, PS 5/6 C, PS 5/6 PK, PS 4/5, and PG 8/1 had lesser siderophore production after FeCl addition, AT 7/1 C, AT 7/1 K, and KI 8/1 also showed good siderophore production. These findings demonstrated that different rhizobacteria isolates had varying capacities for producing siderophores, which are crucial for boosting plant iron availability and promoting growth and resistance to pathogens.
In terms of their capacity to fix atmospheric nitrogen, AT 7/1 C, BMS 8/3 P, KI 8/1, PS 6/1, PS 6/5, PT 7/2, PT 8/1, PT 8/2, and TT 8/3 CR all showed positive nitrogen fixation activity. By supplying vital nitrogen, these isolates may increase soil fertility and promote plant growth. However, CL 4/1, PG 7/3 CK, KI 8/3, KI 8/4, LB 4/1 C, LB 5/1 K, PG 7/3 CK, PG 9/2 C, PS 5/6 C, and PS 5/6 PK did not show significant nitrogen fixation activity, suggesting the variation in nitrogen fixation potential among the tested isolates. These results showed that rhizobacteria isolate with good nitrogen fixation ability could be used as biofertilizers to enhance agricultural productivity and reduce dependence on chemical fertilizers.
The ability to produce HCN showed that most isolates showed variation in HCN production. Based on the results, BMS 8/3 P, PS 5/6 C, and TT 8/3 CR showed the best HCN production, while CL 4/1, PG 7/3 CK, and PG 9/2 C had no significant potential. Some isolates, such as AT 7/1 C, KI 8/1, and PS 4/2, showed positive HCN production, which could contribute to plant protection against pathogens by producing toxic compounds. But HCN production was absent from CL 8/1, KI 8/3, LB 4/1 C, LB 4/2, PG 7/2, PG 7/3 CK, PG 8/1, PG 9/2 C, and PT 8/1. These findings showed that rhizobacteria isolates with a high capacity for producing HCN might act as bioprotectants, helping plants fend against pathogen invasions. However, the possible effects of HCN production on the environment and plant health must be taken into account.
Isolates such as AT 7/1 C, BMS 8/3 P, BMS 8/5 H, KI 8/4, LB 4/1 C, LB 4/2, LB 5/1 K, PG 8/1, PG 9/2 C, PS 6/3 A, PT 8/1, PT 8/2, TT 8/3 CR, and TT 8/4 P showed high ACC-deaminase production. This could help reduce ethylene levels in plants, alleviating stress caused by environmental stressors. Other isolates such as PS 5/6 C and PT 7/2 did not show significant ACC-deaminase production. These results suggested that rhizobacteria isolates with good ACC-deaminase production could function as biofertilizers, assisting plants in maintaining growth and productivity under stress conditions, as well as improving overall plant health.
The ability to produce HCN showed that most isolates showed variation in HCN production. Based on the results, BMS 8/3 P, PS 5/6 C, and TT 8/3 CR showed the best HCN production, while CL 4/1, PG 7/3 CK, and PG 9/2 C had no significant potential. Some isolates, such as AT 7/1 C, KI 8/1, and PS 4/2, showed positive HCN production, which could contribute to plant protection against pathogens by producing toxic compounds. However, CL 8/1, KI 8/3, LB 4/1 C, LB 4/2, PG 7/2, PG 7/3 CK, PG 8/1, PG 9/2 C, and PT 8/1 did not show HCN production. These results suggested that rhizobacteria isolates with good HCN production potential could serve as bioprotectants, facilitating plants to defend against pathogen attacks. However, there is a need to consider the potential environmental and plant health impacts of HCN production.
Isolates such as AT 7/1 C, BMS 8/3 P, BMS 8/5 H, KI 8/4, LB 4/1 C, LB 4/2, LB 5/1 K, PG 8/1, PG 9/2 C, PS 6/3 A, PT 8/1, PT 8/2, TT 8/3 CR, and TT 8/4 P showed high ACC-deaminase production. This could help reduce ethylene levels in plants, alleviating stress caused by environmental stressors. Other isolates such as PS 5/6 C and PT 7/2 did not show significant ACC-deaminase production. These results suggested that rhizobacteria isolates with good ACC-deaminase production could function as biofertilizers, assisting plants in maintaining growth and productivity under stress conditions, as well as improving overall plant health.
3.2 Analysis of soil nutrient content before and after using rhizobacteria
The analysis is one of the observations to determine soil characteristics. Soil analysis in the study was carried out twice at the beginning before the application of rhizobacteria and after the application of rhizobacteria which are shown in Tables 2 and 3.
Table 2. Soil fertility analysis before the application of rhizobacteria isolates
|
Soil Parameters |
Description |
|
|
Soil Texture |
Sandy loam |
|
|
Sand (%) |
41.17 |
|
|
Dust (%) |
32.46 |
|
|
Clay (%) |
26.37 |
|
|
Soil Reaction |
|
|
|
C-Organic (%) |
0.23 |
Very low |
|
N-Total (%) |
0.01 |
Very low |
|
C/N |
26.92 |
Very high |
|
P2O5 (mg 100/g) |
37.48 |
Medium |
|
K-Available (ppm) |
7.23 |
Very low |
|
K- Exchangeable |
0.39 |
Medium |
|
Na-Exchangeable |
0.11 |
Very low |
|
Ca-Exchangeable |
0.23 |
Very low |
|
Mg-Exchangeable |
0.14 |
Very low |
|
CEC (meq/100g) |
11.90 |
Low |
|
pH H2O |
5.69 |
Slightly acidic |
Source: Testing Service Laboratory, Agricultural Instrument Standardization Application Center, Aceh Province
In Table 2, the soil analysis criteria before the application of rhizobacteria isolates showed that the texture used as the planting medium was clayey with a sand percentage of 41.17%, dust 32.46%, and clay 26.37%. The organic carbon content was very low, total nitrogen was in the very low category, C/N ratio in the very high category, and P2O5 content in the moderate category. Available potassium, exchangeable sodium, calcium, and magnesium were in the very low category. Meanwhile, exchangeable potassium was in the moderate category, cation exchange capacity (CEC) was low, and soil pH (H2O) was slightly acidic.
Table 3 showed that the soil treated with rhizobacteria had an organic carbon content, C/N ratio, exchangeable magnesium, and CEC within the moderate criteria. Total nitrogen, exchangeable sodium, and available potassium were within the low criteria. Exchangeable potassium was high, exchangeable calcium was extremely low, the pH of the soil was within the somewhat acidic range, and the P2O5 level met very high standards. The P2O5 level increased significantly from moderate to very high, according to the data. A similar trend was observed in the increase in C-Organic content from initially very low to moderate conditions. An increase was also observed in the exchangeable K content from moderate to high condition, while CEC rose from initially low to moderate condition.
Table 3. Soil fertility analysis after the application of rhizobacteria isolates
|
Soil Parameter |
Description |
|
|
C-Organic (%) |
2.31 |
Medium |
|
N-Total (%) |
0.18 |
Low |
|
C/N |
12.87 |
Medium |
|
P2O5 (mg 100/g) |
131.71 |
Very high |
|
K-Available (ppm) |
12.56 |
Low |
|
K- Exchangeable |
0.85 |
High |
|
Na-Exchangeable |
0.32 |
Low |
|
Ca-Exchangeable |
0.45 |
Very low |
|
Mg-Exchangeable |
1.97 |
Medium |
|
CEC (meq/100g) |
17.09 |
Medium |
|
pH H2O |
6.54 |
Slightly acidic |
Source: Testing Service Laboratory, Agricultural Instrument Standardization Application Center, Aceh Province
3.3 Agronomic testing of rhizobacteria on the growth of Aceh patchouli (Pogostemon cablin Benth.)
The results of testing various potential rhizobacteria isolates on patchouli plant growth, including parameters such as plant height, number of branches and leaves, root volume, plant wet biomass, and plant dry biomass, are shown in Table 4 and Figure 2.
Table 4. Testing of several potential rhizobacteria on the growth of three patchouli varieties
|
Parameters |
Isolation of Rhizobacteria |
Varieties |
||
|
Tapaktuan |
Lhokseumawe |
Sidikalang |
||
|
Plant Height at 12 Weeks After Application (WAA) |
Control |
75.57 ± 3.32Ca |
68.38 ± 6.20Ab |
68.98 ± 10.99Ac |
|
AT 7/1 C |
85.00 ± 7.21Ea |
93.00 ± 3.12Gb |
102.00 ± 5.89Jc |
|
|
BMS 8/3 P |
89.67 ± 5.13Ga |
89.16 ± 4.64Fa |
99.16 ± 4.19Ib |
|
|
CL 4/1 |
72.01 ± 6.92Ba |
80.86 ± 3.80Db |
81.70 ± 2.66Cc |
|
|
KI 8/1 |
79.48 ± 6.83Da |
86.91 ± 6.53Ec |
84.18 ± 10.92Db |
|
|
KI 8/4 |
70.33 ± 4.50ABb |
68.67 ± 5.00Aa |
81.33 ± 3.05Cc |
|
|
LB 4/2 |
92.33 ± 3.05Ha |
92.33 ± 1.52Ga |
93.89 ± 1.16Gb |
|
|
PG 7/3 CK |
88.00 ± 1.50FGa |
96.33 ± 2.51Ic |
90.67 ± 1.25Fb |
|
|
PG 9/2 C |
103.00 ± 3.04Ja |
102.67 ± 2.36Ja |
106.17 ± 2.92Kb |
|
|
PS 4/2 |
86.45 ± 4.01EFa |
72.90 ± 0.60Ba |
73.20 ± 9.33Bb |
|
|
PS 4/5 |
69.58 ± 8.12Aa |
77.33 ± 5.13Cb |
85.67 ± 3.05DEc |
|
|
PS 6/3 A |
74.21 ± 7.65Cb |
79.70 ± 8.99Da |
74.45 ± 7.17Ba |
|
|
PS 5/6 PK |
70.78 ± 4.52ABa |
77.67 ± 4.72Cc |
86.33 ± 2.51Eb |
|
|
PT 8/2 |
92.33 ± 0.57Hb |
95.67 ± 6.42Ha |
101.44 ± 3.35Jc |
|
|
TT 8/4 P |
97.33 ± 4.04Ia |
96.83 ± 4.93Ia |
96.83 ± 2.75Ha |
|
|
Number of Branches in 12 WAA |
Control |
19.67 ± 3.05Ba |
25.83 ± 1.89Ec |
20.50 ± 1.32ABb |
|
AT 7/1 C |
17.67 ± 1.52Aa |
18.16 ± 1.04Bb |
25.67 ± 9.07Cc |
|
|
BMS 8/3 P |
18.33 ± 2.51Aa |
21.67 ± 3.51Db |
21.33 ± 2.25Bb |
|
|
CL 4/1 |
37.00 ± 8.04Hc |
28.33 ± 7.25Fa |
31.90 ± 3.90DEb |
|
|
KI 8/1 |
24.16 ± 2.02DEa |
33.00 ± 11.13Ic |
31.17 ± 5.10DEb |
|
|
KI 8/4 |
23.33 ± 1.52Da |
25.33 ± 1.52Eb |
32.00 ± 1.00Ec |
|
|
LB 4/2 |
30.67 ± 1.52Fa |
36.77 ± 3.28Jb |
38.96 ± 4.50Fc |
|
|
PG 7/3 CK |
38.50 ± 1.50Ib |
38.50 ± 0.50Kb |
37.83 ± 0.76Fa |
|
|
PG 9/2 C |
36.50 ± 1.50Hc |
31.00 ± 1.00Hb |
30.16 ± 1.75Da |
|
|
PS 4/2 |
20.67 ± 6.65BCc |
19.16 ± 4.19Ba |
21.00 ± 3.12ABb |
|
|
PS 4/5 |
25.33 ± 2.08Ea |
25.00 ± 1.00Ea |
26.67 ± 0.57Cb |
|
|
PS 6/3 A |
25.16 ± 3.88Ea |
29.00 ± 3.50FGb |
30.33 ± 3.01Dc |
|
|
PS 5/6 PK |
21.33 ± 2.08Cb |
20.33 ± 2.08Ca |
25.67 ± 1.52Cc |
|
|
PT 8/2 |
32.00 ± 1.00Gb |
29.67 ± 2.08Ga |
38.55 ± 2.91Fc |
|
|
Number of Leaves in 12 WAA |
Control |
177.83 ± 31.48Ba |
198.33 ± 35.26BCDb |
193.33 ± 12.05Ab |
|
AT 7/1 C |
198.67 ± 29.70DEa |
194.16 ± 54.70BCa |
307.67 ± 62.09Jb |
|
|
BMS 8/3 P |
204.00 ± 44.44DEa |
205.83 ± 56.78CDEa |
218.16 ± 43.41CDb |
|
|
CL 4/1 |
289.33 ± 52.77Gc |
272.50 ± 79.54Gb |
234.00 ± 20.66EFa |
|
|
KI 8/1 |
210.33 ± 18.87Ea |
296.33 ± 99.38Hc |
242.83 ± 21.50FGb |
|
|
KI 8/4 |
229.33 ± 13.79Fa |
236.67 ± 5.68Fb |
259.33 ± 7.57Hc |
|
|
LB 4/2 |
180.00 ± 10.44Ba |
191.00 ± 30.11Bb |
275.46 ± 48.24Ic |
|
|
PG 7/3 CK |
184.67 ± 11.54DCc |
189.00 ± 0.86Aba |
193.83 ± 3.40Ab |
|
|
PG 9/2 C |
337.67 ± 12.88Hb |
326.16 ± 2.75Ia |
361.33 ± 5.85Kc |
|
|
PS 4/2 |
182.00 ± 3.00BCc |
227.16 ± 36.19Fb |
219.67 ± 36.68CDa |
|
|
PS 4/5 |
199.33 ± 22.23DEa |
212.00 ± 45.90Ea |
224.00 ± 37.16DEb |
|
|
PS 6/3 A |
142.33 ± 11.62Aa |
337.83 ± 61.72Ic |
258.16 ± 72.47GHb |
|
|
PS 5/6 PK |
192.33 ± 3.05CDa |
209.33 ± 22.27DEb |
195.33 ± 0.57ABa |
|
|
PT 8/2 |
192.67 ± 6.80CDa |
197.67 ± 8.50BCDa |
207.11 ± 2.22BCb |
|
|
TT 8/4 P |
142.33 ± 20.25Aa |
177.83 ± 12.29Ab |
195.67 ± 65.54ABc |
|
|
Root Volume |
Control |
45.83 ± 10.07Aa |
62.50 ±7.07Cc |
52.50 ± 18.71Ab |
|
AT 7/1 C |
49.67 ± 4.78ABa |
50.00 ± 5.40Aa |
70.83 ± 2.36Eb |
|
|
BMS 8/3 P |
57.33 ± 4.19CDa |
58.33 ± 1.18BCa |
67.50 ± 3.54CDEb |
|
|
CL 4/1 |
81.67 ± 20.95Fb |
114.17 ± 45.15Hc |
70.00 ± 15.41DEa |
|
|
KI 8/1 |
89.17 ± 11.96Gb |
90.83 ± 26.01Fb |
83.33 ± 8.50Fa |
|
|
KI 8/4 |
66.33 ± 4.19Ea |
71.00 ± 2.45Db |
71.67 ± 2.62Eb |
|
|
LB 4/2 |
53.33 ± 4.19BCa |
56.67 ± 5.25Bb |
70.55 ± 1.97Ec |
|
|
PG 7/3 CK |
65.83 ± 1.18Eb |
71.67 ± 1.18Dc |
63.33 ± 3.12Ca |
|
|
PG 9/2 C |
85.00 ± 2.04FGc |
50.83 ± 1.18Aa |
57.50 ± 2.04Bb |
|
|
PS 4/2 |
101.67 ± 23.21Hc |
85.00 ± 14.28Eb |
65.00 ± 22.73CDa |
|
|
PS 4/5 |
60.00 ± 4.96DEc |
56.33 ± 2.49Bb |
53.00 ± 7.78ABa |
|
|
PS 6/3 A |
65.00 ± 19.47Ea |
100.83 ± 12.47Gb |
65.83 ± 12.47CDa |
|
|
PS 5/6 PK |
69.33 ± 5.24Eb |
59.67 ± 3.29BCa |
66.33 ± 2.49CDEa |
|
|
PT 8/2 |
82.00 ± 3.26Fa |
95.00 ± 4.89Fb |
102.44 ± 2.347Gc |
|
|
TT 8/4 P |
57.50 ± 3.30CDa |
57.50 ± 2.04Ba |
67.50 ± 5.40CDEb |
|
|
Wet Biomass Weight of Plants |
Control |
228.33 ± 47.05Bc |
224.16 ± 25.16Ab |
197.50 ± 18.87Aa |
|
AT 7/1 C |
370.00 ± 62.44Ha |
360.00 ± 65.57EFa |
468.33 ± 63.31Hb |
|
|
BMS 8/3 P |
321.33 ± 34.64Fa |
418.33 ± 100.66Hc |
326.67 ± 11.54Eb |
|
|
CL 4/1 |
338.16 ± 97.20FGb |
348.33 ± 135.30CDEc |
255.83 ± 23.62BCa |
|
|
KI 8/1 |
248.33 ± 35.02BCa |
375.83 ± 213.69FGc |
289.16 ± 26.73Db |
|
|
KI 8/4 |
228.00 ± 15.39Ba |
256.67 ± 7.23Bb |
267.33 ± 10.06BCDc |
|
|
LB 4/2 |
280.00 ± 10.44DEa |
324.33 ± 40.50CDb |
380.00 ± 13.33Fc |
|
|
PG 7/3 CK |
353.00 ± 6.55GHc |
351.00 ± 40.50DEFa |
363.50 ± 13.33Fb |
|
|
PG 9/2 C |
360.00 ± 5.00GHb |
321.67 ± 9.96 Ca |
355.00 ± 3.04 Fb |
|
|
PS 4/2 |
324.17 ± 71.07Fc |
260.83 ± 2.88Bb |
249.83 ± 8.66Ba |
|
|
PS 4/5 |
235.33 ± 11.67Ba |
278.67 ± 39.23Bb |
257.33 ± 37.55BCa |
|
|
PS 6/3 A |
170.83 ± 35.02Aa |
390.00 ± 11.84Gc |
279.16 ± 34.84CDb |
|
|
PS 5/6 PK |
292.33 ±3.05Ea |
276.00 ± 125.79Bb |
285.33 ± 85.67Dab |
|
|
PT 8/2 |
359.33 ± 56.09GHb |
331.00 ± 35.53CDa |
431.11 ± 17.61Gc |
|
|
TT 8/4 P |
260.67 ± 51.62CDa |
265.00 ± 43.58Ba |
435.00 ± 148.49 Db |
|
|
Dry Biomass Weight of Plants |
Control |
43.50 ± 2.50BCa |
38.17 ± 1.04Ab |
33.83 ± 10.02Ac |
|
AT 7/1 C |
96.83 ± 12.35Ja |
97.50 ± 13.92Ha |
122.50 ± 10.90Ib |
|
|
BMS 8/3 P |
98.33 ± 15.28Jb |
103.33 ± 24.28Ic |
83.33 ± 8.78Ea |
|
|
CL 4/1 |
57.00 ± 6.24Eb |
65.17 ± 10.37Dc |
48.33 ± 8.08Ca |
|
|
KI 8/1 |
49.00 ± 7.94Da |
70.50 ± 27.99Ec |
51.83 ± 14.84Cb |
|
|
KI 8/4 |
47.67 ± 6.43CDa |
54.33 ± 8.02Cc |
50.33 ± 12.42Cb |
|
|
LB 4/2 |
73.33 ± 12.58Ga |
81.67 ± 15.28Fb |
96.67 ± 22.55Fc |
|
|
PG 7/3 CK |
87.33 ± 2.47Ib |
104.67 ± 2.08Ic |
84.17 ± 1.44Ea |
|
|
PG 9/2 C |
118.33 ± 2.89Ka |
130.83 ± 3.82Jb |
143.33 ± 9.46Jc |
|
|
PS 4/2 |
78.17 ± 21.10GHc |
40.50 ± 6.00Aa |
42.67 ± 9.61Bb |
|
|
PS 4/5 |
38.67 ± 5.51Aa |
47.67 ± 7.37Bb |
56.67 ± 4.73Dc |
|
|
PS 6/3 A |
39.33 ± 9.57ABa |
57.83 ± 9.93Cc |
49.83 ± 21.00Cb |
|
|
PS 5/6 PK |
44.00 ± 7.21Ca |
53.67 ± 8.50Cc |
50.67 ± 5.13Cb |
|
|
PT 8/2 |
78.33 ± 5.77Hb |
75.00 ± 10.00Ea |
108.87 ± 26.94Hc |
|
|
TT 8/4 P |
93.50 ± 12.02Fa |
90.00 ± 15.21Gb |
108.33 ± 16.27Gc |
|
Note: Duncan's Multiple Range Test was used to assess the data (p > 0.05). Lowercase characters are read vertically (columns) and capital letters horizontally (rows). No discernible change is indicated by identical notation.
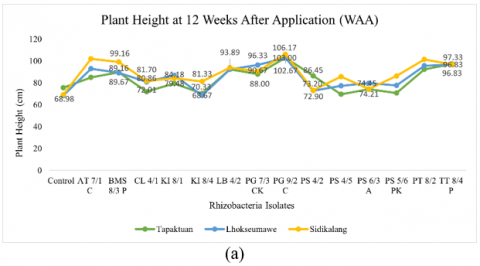
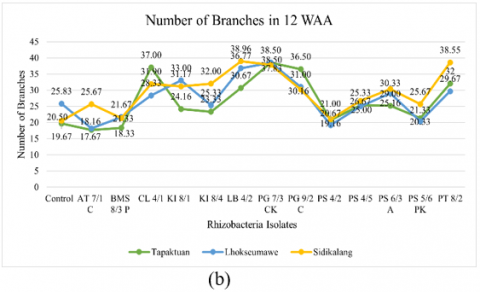
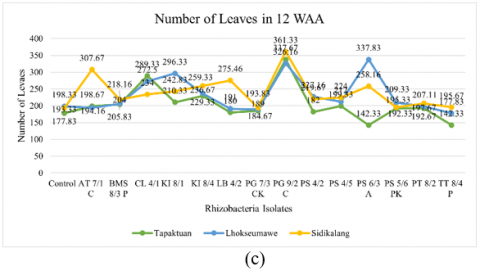
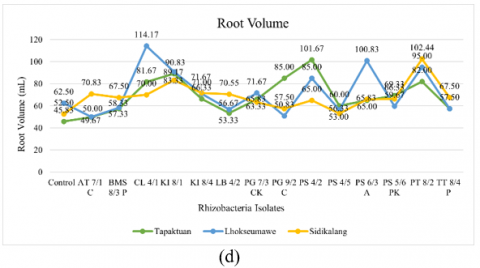
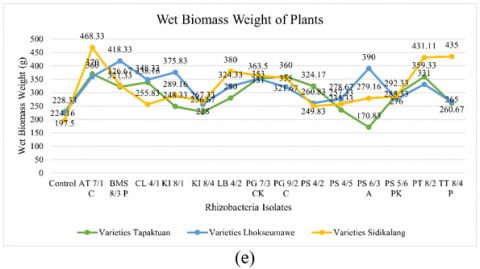
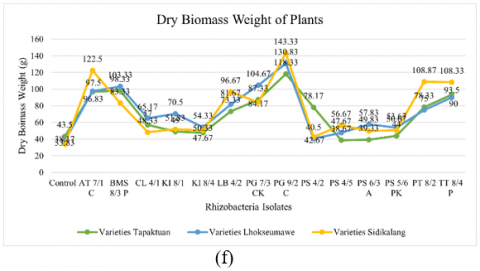
Figure 2. Testing of several potential rhizobacteria on the growth of three patchouli varieties (a) Plant height, (b) Number of branches, (c) Number of leaves, (d) Root volume, (e) Wet biomass weight, and (e) Dry biomass weight
A significant substantial interaction effect on patchouli plant growth was seen when many rhizobacteria isolates were applied to various types (Figure 3). Considering the group that was tested, nine rhizobacteria isolates significantly affected plant height compared to each variety, as shown in Table 4. PG 9/2 C produced the highest plant height at 106.17 cm in the Sidikalang variety, followed by AT 7/1 C (102.00 cm), PT 8/2 (101.44 cm), and BMS 8/3 P (99.16 cm). Other isolates such as LB 4/2 reached a height of 93.89 cm, PG 7/3 (90.67 cm), PS 5/6 PK (86.33 cm), CL 4/1 (81.70 cm), and KI 8/4 (81.33 cm). It was discovered that these isolates worked well to increase the Sidikalang variety of patchouli plants' height development. The rhizobacteria isolate PG 7/3 CK showed the best plant height in the Lhokseumawe variety compared to others, with a height of 96.33 cm, followed by KI 8/1 (86.91 cm), and PS 6/3 A (79.70 cm). Meanwhile, PS 4/2 produced the best plant height in the Tapaktuan variety compared to others, with a height of 86.45 cm, while the control treatment had 75.57 cm.
Figure 3. Comparison root volume of patchouli plants between control treatment and application of rhizobacteria isolate PG 9/2 C on varieties (a) Sidikalang (b) Lhokseumawe and (c) Tapak Tuan
The number of branches in patchouli plants was significantly affected by rhizobacteria isolates, 12 isolates had significantly different branch numbers. Furthermore, within the Sidikalang variety, the highest number of branches was observed in AT 7/1 C, KI 8/4, LB 4/2, PS 4/2, PS 4/5, PS 6/3 A, PS 5/6 PK, PT 8/2, and TT 8/4. CL 4/1, PG 9/2 C, and PG 7/3 CK had the highest number of branches in the Tapaktuan variety, while BMS 8/3 P, KI 8/1, PG 7/3 CK, and the control treatment had the highest number of branches in the Lhokseumawe variety. The results showed that rhizobacterial isolates positively influenced the branch number of patchouli plants. LB 4/2, PG 7/3 CK, and PG 9/2 C performed well, showing relatively high branch numbers across all varieties. In almost all varieties, PG 7/3 CK produced the most branches, with 38.50 in the Tapaktuan and Lhokseumawe varieties and 37.83 in Sidikalang. These findings demonstrated that rhizobacteria isolates could improve plant growth by increasing branch numbers.
Based on observations, there were eight rhizobacteria isolates that significantly affected the root volume in patchouli plants. The isolates PG 9/2 C, PS 4/2, PS 4/5, and PS 5/6 PK showed the highest root volume in the Tapaktuan variety. CL 4/1, KI 8/1, PG 7/3 CK, PS 6/3 A, and the control showed the highest root volumes in the Lhokseumawe variety, while AT 7/1, BMS 8/3 P, KI 8/4, LB 4/2, PT 8/2, and TT 8/4 P produced the highest root volumes in the Sidikalang variety. Among the varieties, isolates PS 4/5 did not significantly differ from isolates PS 6/3 A and TT 8/4 P. Overall, rhizobacterial isolates had a positive effect on increasing root volume in patchouli plants. Isolates such as PS 4/2, CL 4/1, and PT 8/2 were the most effective in supporting root growth compared to the control. PS 4/2 produced the highest root volume in the Tapaktuan variety at 101.67 mL, CL 4/1 had the highest root volume in Lhokseumawe at 114.17 mL, and PT 8/2 had the highest root volume in Sidikalang at 102.44 mL. Root volume variation was observed among the different varieties, with Sidikalang showing higher volume compared to others, as shown in Figure 3.
The wet biomass weight of plants varied significantly among the rhizobacteria isolates and varieties, as Table 4 illustrates. Based on observation, 10 treatments with rhizobacteria isolates showed significantly different wet biomass weight in the patchouli plants. Isolates AT 7/1 C, PG 9/2 C, PS 4/2, PS 5/6 PK, and the control showed the highest wet biomass weight in the Tapaktuan variety. In Lhokseumawe variety, BMS 8/3 P, CL 4/1, KI 8/1, PS 4/5, and PS 6/3 A produced the highest wet biomass weight, while KI 8/4, LB 4/2, PG 7/3 CK, PT 8/2, and TT 8/4 P were found in Sidikalang Isolates AT 7/1 C, BMS 8/3 P, and PG 9/2 C performed better than the control in promoting biomass growth. The findings indicated that, among all kinds, AT 7/1 C had the maximum wet biomass, with 370.00 g in Tapaktuan, 360.00 g in Lhokseumawe, and 468.33 g in Sidikalang. The control showed the lowest wet biomass weight in all varieties, while the Sidikalang variety had the highest for certain isolates.
Rhizobacteria isolates positively influenced the increase in dry biomass weight of patchouli plants. Based on the results, AT 7/1 C, LB 4/2, PG 9/2 C, PS 4/5, PT 8/2, and TT 8/4 P showed the highest dry biomass weight in the Sidikalang variety. In Lhokseumawe, BMS 8/3 P, CL 4/1, KI 8/1, PG 7/3 CK, PS 6/3 A, and PS 5/6 PK showed the highest. In the meantime, the Tapaktuan variety's maximum dry biomass weight was displayed by isolates PS 4/2 and the control. The dry biomass weight varied significantly among rhizobacterial isolates and varieties. In comparison to the control, isolates PG 9/2 C, AT 7/1 C, and BMS 8/3 P were the most successful in increasing dry biomass weight. Furthermore, PG 9/2 C showed the highest dry biomass weight across all varieties, with 118.33 g in Tapaktuan, 130.83 g in Lhokseumawe, and 143.33 g in Sidikalang. In comparison, the control had the lowest dry biomass weight across all varieties.
3.4 Molecular identification
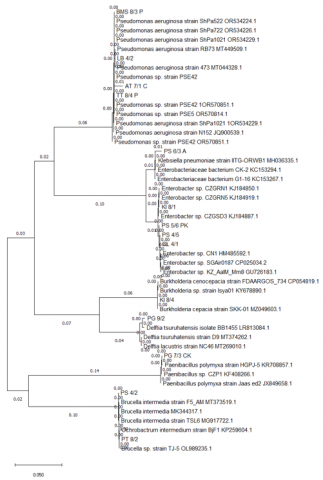
Several rhizobacterial isolates were from the same species but different strains, according to the molecular identification results in Table 5. Pseudomonas aeruginosa, Enterobacter sp., Brucella intermedia, Burkholderia cepacia, Delftia tsuruhatensis, and several bacteria from the families Paenibacillus and Klebsiella were found to be the majority of the isolates. Some isolates including BMS 8/3 P, LB 4/2, and TT 8/4 P showed 100% similarity with specific strains of Pseudomonas aeruginosa, while AT 7/1 C had 99.12%. These suggested isolates were Pseudomonas aeruginosa and have strong potential as rhizobacteria agents for plant growth. Isolates PS 4/2 and PT 8/2 were identified as Brucella intermedia, while CL 4/1, PS 4/5, and PS 5/6 PK were classified as Enterobacter sp., strain SGAir0187. Moreover, isolate KI 8/1 was identified as a different strain, Enterobacter sp., strain CZGRN1. Isolate PG 9/2 C was identified as Delftia tsuruhatensis, while Paenibacillus polymyxa, strain HGPJ-5, was identified in PG 7/3 CK. PS 6/3 A was identified as Klebsiella pneumoniae, strain IITG-ORWB1, and KI 8/4 was identified as Burkholderia cepacian, strain SKK-01. The phylogenetic tree in Figure 4 illustrates the evolutionary relationships of these isolates.
Table 5. Sequencing results of the 16S rRNA gene of rhizobacteria isolates from the patchouli rhizosphere in Aceh according to data available in NCBI (BLASTX)
|
Isolate Code |
Description |
Max Score |
Total Score |
Quary Cover |
E Value |
Identity |
Access Number |
|
AT 7/1 C |
Pseudomonas aeruginosa strain N152 |
2447 |
2447 |
100% |
0.0 |
99.12% |
JQ900539.1 |
|
Pseudomonas aeruginosa strain ShPa1021 |
2442 |
2442 |
100% |
0.0 |
99.04% |
OR534229.1 |
|
|
Pseudomonas aeruginosa strain ShPa722 |
2442 |
2442 |
100% |
0.0 |
99.04% |
OR534226.1 |
|
|
BMS 8/3 P |
Pseudomonas aeruginosa strain ShPa1021 |
2580 |
2580 |
100% |
0.0 |
100.00% |
OR534229.1 |
|
Pseudomonas aeruginosa strain ShPa722 |
2580 |
2580 |
100% |
0.0 |
100.00% |
OR534226.1 |
|
|
Pseudomonas aeruginosa strain ShPa522 |
2580 |
2580 |
100% |
0.0 |
100.00% |
OR534224.1 |
|
|
CL 4/1 |
Enterobacter sp. SGAir0187 |
2571 |
20412 |
100% |
0.0 |
99.86% |
CP025034.2 |
|
Enterobacter sp. KZ AaIM Mm8 |
2571 |
2571 |
100% |
0.0 |
99.86% |
GU726183.1 |
|
|
Enterobacter sp. CN1 |
2566 |
2566 |
100% |
0.0 |
99.79% |
HM485592.1 |
|
|
KI 8/1 |
Enterobacter sp. CZGRN1 |
2567 |
2567 |
100% |
0.0 |
99.79% |
KJ184850.1 |
|
Enterobacter sp. CZGRN5 |
2562 |
2562 |
100% |
0.0 |
99.71% |
KJ184919.1 |
|
|
Enterobacter sp. CZGSD3 |
2562 |
2562 |
100% |
0.0 |
99.71% |
KJ184887.1 |
|
|
KI 8/4 |
Bulkholderia cepacia strain SKK-01 |
2579 |
2579 |
100% |
0.0 |
100% |
MZ049603.1 |
|
Bulkholderia cenocepacia strain FDAARGOS 734 |
2573 |
2573 |
100% |
0.0 |
99.93% |
CP054819.1 |
|
|
Bulkholderia sp. strain Isya01 |
2573 |
2573 |
100% |
0.0 |
99.93% |
KY678890.1 |
|
|
LB 4/2 |
Pseudomonas aeruginosa strain RB73 |
2538 |
2538 |
100% |
0.0 |
100.00% |
MT449509.1 |
|
Pseudomonas aeruginosa strain 473 |
2538 |
2538 |
100% |
0.0 |
100.00% |
MT044328.1 |
|
|
Pseudomonas aeruginosa strain ShPa1021 |
2538 |
2532 |
100% |
0.0 |
100.00% |
OR534229.1 |
|
|
PG 7/3 CK |
Paenbacillus polymyxa strain HGPJ-5 |
2604 |
2604 |
100% |
0.0 |
99.86% |
KR708857.1 |
|
Paenbacillus sp. CZP1 |
2604 |
2604 |
100% |
0.0 |
99.86% |
KF408266.1 |
|
|
Paenbacillus polymyxa strain Jaas ed2 |
2604 |
2604 |
100% |
0.0 |
99.86% |
JX849658.1 |
|
|
PG 9/2 C |
Delftia tsuruhatensis strain BB1455 |
2475 |
12378 |
100% |
0.0 |
99.49% |
LR813084.1 |
|
Delftia tsuruhatensis strain D9 |
2464 |
2464 |
100% |
0.0 |
99.34% |
MT374262.1 |
|
|
Delftia lacustris strain NC46 |
2464 |
2464 |
100% |
0.0 |
99.34% |
MT269010.1 |
|
|
PS 4/2 |
Brucella intermedia strain F5 |
2481 |
2481 |
100% |
0.0 |
99.93% |
MT373519.1 |
|
Brucella intermedia |
2481 |
2481 |
100% |
0.0 |
99.93% |
MK344317.1 |
|
|
Brucella intermedia strain TSL6 |
2481 |
2481 |
100% |
0.0 |
99.93% |
MG917722.1 |
|
|
Orchrobactrum intermedium strain BjF1 |
2481 |
2481 |
100% |
0.0 |
99.93% |
KR269740.1 |
|
|
PS 4/5 |
Enterobacter sp. SGAir0187 |
2564 |
20353 |
100% |
0.0 |
99.86% |
CP025034.2 |
|
Enterobacter sp. KZ AaIM Mm8 |
2564 |
2564 |
100% |
0.0 |
99.86% |
GU726183.1 |
|
|
Enterobacter sp. CN1 |
2558 |
2558 |
100% |
0.0 |
99.78% |
HM485592.1 |
|
|
PS 6/3 A |
Klebsiella pneumonia strain IITG-ORWB1 |
2523 |
2523 |
99% |
0.0 |
99.80% |
MH036335.1 |
|
Enterobacteriaceae bacterium CK-2 |
2523 |
2523 |
99% |
0.0 |
99.80% |
KC153294.1 |
|
|
Enterobacteriaceae bacterium G1-16 |
2523 |
2523 |
99% |
0.0 |
99.80% |
KC153267.1 |
|
|
PS 5/6 PK |
Enterobacter sp. SGAir0187 |
2558 |
20325 |
100% |
0.0 |
99.64% |
CP025034.2 |
|
Enterobacter sp. KZ AaIM |
2558 |
2558 |
100% |
0.0 |
99.64% |
GU726183.1 |
|
|
Enterobacter sp. CZGSD3 |
2553 |
2553 |
100% |
0.0 |
99.57% |
KJ184887.1 |
|
|
PT 8/2 |
Brucella intermedia strain TSL6 |
2494 |
2494 |
100% |
0.0 |
100.00% |
MG917722.1 |
|
Ochrobactrum intermedium strain BjF1 |
2494 |
2494 |
100% |
0.0 |
100.00% |
KP259604.1 |
|
|
Brucella sp. strain TJ-5 |
2494 |
2494 |
100% |
0.0 |
100.00% |
OL989235.1 |
|
|
TT 8/4 P |
Pseudomonas aeruginosa strain ShPa1021 |
2532 |
2532 |
100% |
0.0 |
100.00% |
OR534229.1 |
|
Pseudomonas aeruginosa strain ShPa722 |
2532 |
2532 |
100% |
0.0 |
100.00% |
OR534226.1 |
|
|
Pseudomonas aeruginosa strain ShPa522 |
2532 |
2532 |
100% |
0.0 |
100.00% |
OR534224.1 |
Figure 4. Phylogenetic tree of rhizobacterial isolates from the rhizosphere of patchouli (Pogostemon cablin) in Aceh
An auxin hormone called IAA is important for plant growth and development because it promotes tissue differentiation, lateral and adventitious root production, and cell division and elongation. These characteristics help roots and stems grow longer, which improves the plant's capacity to take up nutrients and water from the soil [33, 34]. Numerous factors, including the presence of microorganisms like rhizobacteria, can affect the formation of IAA. Numerous bacteria that live in the rhizosphere of plants produce the phytohormone IAA. These bacteria use a variety of IAA biosynthesis pathways, and some strains have multiple pathways. Study [34] shows that rhizosphere isolates may generate IAA at different doses up to 3.609 mg/L. Study [35] claimed that the soil microbiota influenced plant growth and development and contributed to health. The rhizosphere and roots of Ceanothus velutinus yielded 24 PGPR-producing IAA, which were found to promote plant growth. This finding highlighted how crucial rhizobacteria bacteria are for promoting plant growth by producing vital growth hormones.
Several studies have demonstrated that the application of IAA can stimulate plant growth, leading to increases in height, root length, and other growth parameters [36]. Thus, IAA plays a vital role in promoting physical growth and improving plants' ability to absorb water and nutrients from the soil. For instance, study [37] found that Serratia marcescens strain MBC1 produced IAA, resulting in significant increases in root and stem length in soybean plants. Furthermore, IAA also influenced plant growth in hydroponic systems, as evidenced by its significant effect on celery plants.
IAA production was demonstrated by rhizobacteria isolated from patchouli plants, which improved plant growth and yield [38]. IAA, which is crucial for physiological activities such as lateral root development and stem elongation, was produced by several rhizobacteria isolates in this investigation at different doses. Higher total yields were the result of patchouli plants' vegetative development being promoted by the high concentration that rhizobacteria produced. Bacteria that produce IAA can also increase the productivity of Arabidopsis biomass production and plant propagation [35]. Furthermore, it has been demonstrated that IAA promotes gas exchange and stem diameter expansion, which both improve coffee plant growth [39]. In this study, patchouli plants' stem diameter and shoot and leaf counts were enhanced by the use of IAA-producing rhizobacteria isolates. This demonstrated that the isolates promoted the physical development and general production of the plants.
L-tryptophan is one of the organic substances that naturally occur in plant roots and are utilized by rhizobacteria for IAA production to resist biotic and abiotic stress [36]. Rhizobacteria that produce IAA encourage the host's root system to flourish, which may lessen the requirement for chemical pesticides on patchouli plants. Other studies have shown that rhizobacteria such as Pseudomonas fluorescens, Stenotrophomonas rhizophila, and Bacillus cereus enhance plant growth and resistance to pathogens [40, 41]. According to study [42], IAA-producing rhizobacteria isolates were shown to be effective in inducing plant resistance to viral infections and reducing disease intensity in tobacco plants. This mechanism also applies to patchouli plants, where rhizobacteria compete with pathogens and produce antimicrobial compounds to protect against disease attacks [35]. Enterobacter sp. capable of producing IAA significantly enhances maize growth by increasing root length, plant height, fresh weight, and dry weight [36]. Therefore, the application of IAA-producing rhizobacteria can be an environmentally friendly alternative to reduce dependence on chemical pesticides. The results indicated that all rhizobacterial isolates were capable of producing IAA. The rhizobacteria with the highest IAA production included Enterobacter sp., Delftia tsuruhatensis, and Paenibacillus polymyxa. In contrast, Pseudomonas aeruginosa produced less IAA compared to the other species.
Competition for iron consumption through siderophore synthesis is one of the ways rhizobacteria regulate plant diseases [20]. In the rhizosphere, different Plant Growth-Promoting Bacteria (PGPB) help plants absorb iron which speeds up the physiological and biochemical processes of plants in adverse situations. These bacteria can produce and release siderophores, which improve plant nutrition and growth, and they can also control the bioavailability of iron for plants [43-45]. Numerous kinds of bacteria, including Bacillus, Enterobacter, Klebsiella, Paenibacillus, Pseudomonas, and Streptomyces, have been documented to produce siderophores [46-50]. Pseudomonas aeruginosa and Enterobacter sp. were among the rhizobacteria isolates in this investigation that were able to produce siderophores.
Previous studies reported that siderophore-producing rhizobacteria had significant potential in enhancing plant resistance to pathogens and environmental stress. Study [51] found that rhizobacteria isolates from local rice plants (Kamba) could produce siderophores, effectively inhibiting pathogen growth by binding Fe³⁺ for pathogen development. Additionally, siderophore-producing rhizobacteria can help mitigate heavy metal stress in Panax ginseng, enhancing the plant's resilience to harmful environmental conditions [52]. Some rhizobacterial isolates, such as Glutamicibacter nicotianae and Brevibacillus invocatus, have demonstrated the ability to produce siderophores, significantly promoting maize growth [53]. These isolates contribute to improved nutrient absorption, enhancing leaf quality and plant growth rates. As a result, rhizobacteria that produce siderophores can act as potential biocontrol agents, supporting environmentally friendly and sustainable agricultural practices [51].
Both organic and inorganic forms of phosphate are found in soil naturally; these forms are insoluble or only weakly soluble, which restricts the amount of phosphate that soil organisms can use. In acidic soils, inorganic phosphate minerals are typically bonded as AlPO₄·2H₂O (variscite) and FePO₄·2H₂O (strengite); in alkaline soils, they are typically bound as Ca₃(PO₄)₂ (tricalcium phosphate). Phosphate-solubilizing microbes that generate organic acids with functional carboxyl (-COO⁻) and hydroxyl (-O⁻) groups can aid in the release of phosphate from these bonds. This can raise the amount of soluble phosphate in the soil by forming complexes with cations such Ca₃(PO₄)₂, FePO₄·2H₂O, or AlPO₄·2H₂O [54]. In this study, the rhizobacteria isolates capable of solubilizing phosphate included Pseudomonas aeruginosa.
Various types of bacteria capable of solubilizing phosphate in the soil have been identified, including Burkholderia, Azotobacter, Pseudomonas, Bacillus, Enterobacter, Citrobacter, and Pantoea. Numerous organic acids, including oxalate, succinate, tartrate, citrate, lactate, α-ketoglutarate, acetate, formate, propionate, glycolate, glutamate, glyoxylate, malate, and fumarate, are produced by the metabolism of phosphate-solubilizing bacteria. When it comes to phosphate solubilization, each of these bacteria creates a variety of organic acids in variable quantities and types [55]. It has been demonstrated that phosphate can be efficiently dissolved by phosphate-solubilizing rhizobacteria, including Bacillus species and Pseudomonas species [30]. Furthermore, by producing growth-promoting hormones and having antibiosis actions against pathogens, rhizobacteria improve plant growth and yield [56].
The process of transforming atmospheric nitrogen into a form that plants can absorb, known as nitrogen fixation, promotes growth and raises plant output. Despite being present in the atmosphere in large quantities (about 80%) as N₂, plants are unable to directly absorb nitrogen. This method has not been effective in maximizing nitrogen use efficiency, as a significant amount of nitrogen is lost into the atmosphere and water bodies, leading to negative environmental impacts. Therefore, nitrogen fixation in plants offers a practical solution to decrease the reliance on synthetic nitrogen fertilizers and enhance the sustainability of food production systems [57]. Previous studies reported that bacteria with good nitrogen fixation activity had been found in species such as Pseudomonas aeruginosa and Brucella intermedia.Ara.
PGPR such as Bacillus pumilus S1r1 can be a biological alternative for fixing nitrogen from the atmosphere and delaying remobilization in plants, potentially increasing harvest yields [58]. The nitrogenase enzyme system, consisting of two metalloproteins (iron and molybdenum-iron proteins), plays a significant role in converting dinitrogen (N₂) into ammonia (NH₃) through ATP [59]. Various studies have shown some examples of rhizobacteria capable of nitrogen fixation, including Azospirillum spp., Bradyrhizobium spp., Paenibacillus spp., and Paraburkholderia phymatum [60, 61]. This nitrogen fixation process enhances plant growth and supports sustainable agriculture through improved nitrogen efficiency in the soil.
Through the induction of defensive mechanisms against attacks, HCN contributes positively to plant responses to pathogens. Although it is produced in small amounts during the basic metabolism of plants, HCN can be released in larger quantities after tissue damage in cyanogenic species, becoming an important defense mechanism in plants [62]. Various studies have shown that many rhizobacteria, including strains of Pseudomonas, Bacillus, and Enterobacter, are capable of producing HCN. By producing extracellular lytic enzymes and secondary metabolites, PGPR can successfully manage Vicia faba root rot and wilt illnesses. Subsequent research revealed that 34 out of 104 rhizobacteria isolated from the patchouli rhizosphere produced HCN, which shielded plants from infections and other dangerous creatures [63-66]. Halimursyadah et al. [65] also identified nine HCN-producing bacteria isolates from cassava rhizosphere soil. In this study, the bacteria capable of producing HCN included Pseudomonas aeruginosa, Enterobacter sp., and Brucella intermedia.
ACC-deaminase-producing PGPR plays an essential role as 'stress modulators' by reducing ethylene levels in plants, helping to withstand various stress conditions [67]. In this study, ACC-deaminase-producing bacteria were found in species like Pseudomonas aeruginosa, Enterobacter sp., Delftia tsuruhatensis, and Klebsiella pneumoniae. ACC-deaminase (1-aminocyclopropane-1-carboxylate deaminase) in rhizobacteria can break down ACC, ethylene, especially when plants are under stress, is a precursor that can lead to negative effects. Elevated ethylene concentrations can cause chlorosis, inhibit growth, and may even result in plant death.
Previous studies have demonstrated that ACC-deaminase-producing bacteria, such as Streptomyces venezuelae, can enhance plant growth and salinity tolerance by reducing ethylene levels, reactive oxygen species, and Na+ ions [68]. Similarly, Pseudomonas putida and P. fluorescens have shown benefits for wheat plants under salt stress, leading to increased plant height, root length, biomass, and grain yield. Furthermore, ACC-deaminase enzymes in rhizobacteria have been found to improve germination rates and promote the growth of roots and shoots, helping plants tolerate salt stress [69].
According to Danish et al. [70], salinity tolerance in Camelina sativa was significantly increased by soil inoculation with ACC-deaminase-producing PGPR or the expression of the acdS gene in transgenic plants. According to a study by Heydarian et al. [71], ACC-deaminase-producing bacteria are effective in enhancing plant growth under various stress conditions, including salinity and drought. According to Maulidia [72], phosphate-solubilizing rhizobacteria help dissolve tricalcium phosphate, enhancing availability and supporting the biological efficiency of nitrogen fixation. Study [73] found that seed pre-germination treatments with rhizobacteria effectively increased vegetative growth and chili yield, particularly for species such as Azotobacter sp., Bacillus megaterium, B. coagulans, Flavobacterium sp., and Pseudomonas dimuta, which gave the most significant results.
According to Han et al. [74], the D. tsuruhatensis HR4 strain, which was isolated from the rice rhizosphere, successfully inhibited the growth of a number of plant diseases. This resulted from siderophore production, which lessened soil iron restriction. Furthermore, D. tsuruhatensis MTQ3, which was isolated from the tobacco rhizosphere, is a PGPR that is safe for the environment and can improve plant growth and antibacterial activity [75, 76]. Pseudomonas, Azotobacter, Azospirillum, Acetobacter, and Bacillus are other well-known PGPR. Pseudomonas aeruginosa is known to be one of the most powerful PGPR strains among these. According to Ghadamgahi et al. [77], Pseudomonas aeruginosa promotes plant growth by producing siderophores and solubilizing phosphate, which leads to increased mineral absorption and enhanced lateral root development in host plants.
In general, Plant Growth-Promoting Rhizobacteria (PGPR) are crucial for controlling the balance of plant hormones and forming the rhizosphere's microbial population, both of which contribute to the resilience and growth of plants. PGPR has the ability to increase the synthesis of phytohormones that are involved in root growth, stem elongation, and cell differentiation, including auxins (IAA), gibberellins, and cytokinins [78]. Application of rhizobacterial isolates to patchouli plants increased the number of branches by providing nutrients during the vegetative phase, stimulating cell division, differentiating new shoots, and increasing photosynthetic activity through phosphate solubilization [79].
In terms of PGPR interactions with rhizosphere microbial communities, root exudates produced by plants in response to PGPR colonization can also influence the dynamics of soil microbial communities, increasing the population of beneficial microorganisms, and creating a healthier rhizosphere environment [80]. Through this mechanism, PGPR not only acts as a biofertilizer that increases nutrient availability but also functions as a biostimulant and bioprotectant that strengthens the plant defense system and increases productivity. Additionally, PGPR helps create a more stable microbial community in the rhizosphere by promoting nutrient exchange and producing bioactive compounds that support the growth of symbiotic microorganisms. Some PGPR strains are known to enhance the activity of soil enzymes, such as dehydrogenases and phosphatases, which are involved in the mineralization process and improve nutrient availability for plants. In addition, PGPR can affect the soil microbiome by inducing plant systemic resistance (ISR), a defense mechanism that increases plant resistance to pathogen attack and environmental stress. Through competition for nutrients and space, as well as the generation of hydrolytic enzymes that can break down pathogen cell walls, PGPR may also reduce the number of harmful bacteria [81, 82]. Therefore, PGPR has the potential to be a solution for a more sustainable agricultural system since it not only directly promotes plant development through phytohormone synthesis but also stabilizes the soil ecology by improving microbial interactions in the rhizosphere.
Moreover, this study does not address the influence of greenhouse conditions compared to field conditions on the effectiveness of superior PGPR bacteria in enhancing onion crop yields, nor does it evaluate the long-term stability of different PGPR strains. Consequently, a more detailed and specific analysis is warranted to assess the enduring stability and efficacy of these promising bacterial strains.
The PG 9/2 C isolate (D. tsuruhatensis BB1455) significantly enhanced patchouli growth, increasing plant height by 53.91%, the number of leaves by 89.88%, the number of branches by 85.56%, root volume by 85.47%, fresh biomass by 79.75% and dry biomass by 323.68%, respectively, indicating optimal biomass accumulation. Among the 28 tested isolates, 14 demonstrated potential as PGPR based on their physiological characteristics, namely AT 7/1 C, BMS 8/3 P, CL 4/1, KI 8/1, KI 8/4, LB 4/2, PG 7/3 CK, PG 9/2 C, PS 4/2, PS 4/5, PS 6/3 A, PS 5/6 PK, PT 8/2, and TT 8/4 P. The Tapaktuan, Lhokseumawe, and Sidikalang varieties of patchouli plants exhibited the greatest growth promotion by the PG 9/2 C isolate.
[1] Direktorat Jenderal Perkebunan. (2022). Statistik Perkebunan Unggulan Nasional 2020-2022. https://ditjenbun.pertanian.go.id/.
[2] Zikri, I., Kamaruzzaman, S., Susanti, E. (2021). Study on sustainable agriculture and dimension of needs: A case study patchouli farming in Aceh Jaya. In IOP Conference Series: Earth and Environmental Science, Banda Aceh, Indonesia, p. 012038. IOP https://doi.org/10.1088/1755-1315/667/1/012038
[3] Bappeda Aceh. (2015). Road Map Atsiri Aceh. Banda Aceh. https://bappeda.acehprov.go.id/media/2022.08/3_roadmap_sida_2015_edit_tgl_31-12-20151.pdf.
[4] Kumar, R., Swapnil, P., Meena, M., Selpair, S., Yadav, B.G. (2022). Plant growth-promoting rhizobacteria (PGPR): Approaches to alleviate abiotic stresses for enhancement of growth and development of medicinal plants. Sustainability, 14(23): 15514. https://doi.org/10.3390/su142315514
[5] Hakim, S., Naqqash, T., Nawaz, M.S., Laraib, I., Siddique, M.J., Zia, R., Imran, A. (2021). Rhizosphere engineering with plant growth-promoting microorganisms for agriculture and ecological sustainability. Frontiers in Sustainable Food Systems, 5: 617157. https://doi.org/10.3389/fsufs.2021.617157
[6] Kong, Z., Liu, H. (2022). Modification of rhizosphere microbial communities: a possible mechanism of plant growth promoting rhizobacteria enhancing plant growth and fitness. Frontiers in Plant Science, 13: 920813. https://doi.org/10.3389/fpls.2022.920813
[7] Bhat, B.A., Tariq, L., Nissar, S., Islam, S.T., Islam, S.U., Mangral, Z., Dar, T.U.H. (2022). The role of plant-associated rhizobacteria in plant growth, biocontrol and abiotic stress management. Journal of Applied Microbiology, 133(5): 2717-2741. https://doi.org/10.1111/jam.15796
[8] Mu, D., Chen, L., Hua, G., Pu, L., Tian, Z., Liu, Y., Zhang, G., Tang, J. (2023). Diversity and recruitment strategies of rhizosphere microbial communities by camellia fascicularis, a plant species with extremely small populations in China: Plant recruits special microorganisms to get benefit out of them. Diversity. https://doi.org/10.3390/d15121170
[9] Kuźniar, A., Włodarczyk, K., Jurczyk, S., Maciejewski, R., Wolińska, A. (2023). Ecological diversity of bacterial rhizomicrobiome core during the growth of selected wheat cultivars. Biology, 12(8): 1067. https://doi.org/10.3390/biology12081067
[10] Shi, J., Wang, X., Wang, E. (2023). Mycorrhizal symbiosis in plant growth and stress adaptation: From genes to ecosystems. Annual Review of Plant Biology, 74(1): 569-607. https://doi.org/10.1146/annurev-arplant-061722-090342
[11] Almario, J., Fabiańska, I., Saridis, G., Bucher, M. (2022). Unearthing the plant–microbe quid pro quo in root associations with beneficial fungi. New Phytologist, 234(6): 1967-1976. https://doi.org/10.1111/nph.18061
[12] Dunn, M.F., Becerra-Rivera, V.A. (2023). The biosynthesis and functions of polyamines in the interaction of plant growth-promoting rhizobacteria with plants. Plants, 12(14): 2671. https://doi.org/10.3390/plants12142671
[13] Khabbaz, S.E., Ladhalakshmi, D., Babu, M., Kandan, A., Ramamoorthy, V., Saravanakumar, D., Kandasamy, S. (2019). Plant growth promoting bacteria (PGPB)—A versatile tool for plant health management. Canadian Journal of Pesticides & Pest Management, 1: 1-25. https://doi.org/10.34195/can.j.ppm.2019.05.001
[14] Borah, B., Ahmed, R., Hussain, M., Phukon, P., Wann, S.B., Sarmah, D.K., Bhau, B.S. (2018). Suppression of root-knot disease in Pogostemon cablin caused by Meloidogyne incognita in a rhizobacteria mediated activation of phenylpropanoid pathway. Biological Control, 119: 43-50. https://doi.org/10.1016/j.biocontrol.2018.01.003
[15] Das, P.P., Singh, K.R., Nagpure, G., Mansoori, A., Singh, R.P., Ghazi, I.A., Singh, J. (2022). Plant-soil-microbes: A tripartite interaction for nutrient acquisition and better plant growth for sustainable agricultural practices. Environmental Research, 214: 113821. https://doi.org/10.1016/j.envres.2022.113821
[16] Halimursyadah, H., Fidia, Y., Husna, R. (2024). Effectiveness test of indigenous plant growth promoting rhizobacteria (PGPR) isolate on the growth of Acehnese patchouli (Pogostemon cablin Benth.) Sidikalang variety. Agrin, 28(1): 27-36. https://doi.org/10.20884/1.agrin.2024.28.1.844
[17] Zilaie, M.N., Arani, A.M., Etesami, H., Dinarvand, M., Dolati, A. (2022). Halotolerant plant growth-promoting rhizobacteria-mediated alleviation of salinity and dust stress and improvement of forage yield in the desert halophyte Seidlitzia Rosmarinus. Environmental and Experimental Botany, 201: 104952. https://doi.org/10.1016/j.envexpbot.2022.104952
[18] Saraswati, R., Husen, E., Simanungkalit, R.D.M. (2007). Metode analisis biologi tanah. balai besar penelitian dan pengembangan sumberdaya lahan pertanian. http://balittanah.litbang.deptan.go.id.
[19] Glickman, E., Dessaux, Y. (1995). A critical examination of specificity of the Salkowski reagent for indolic compounds produced by phytopathogenic bacteria. Applied and Environmental Microbiology, 61: 793-796. https://doi.org/10.1128/aem.61.2.793796.1995
[20] Sriwati, R., Maulidia, V., Intan, N., Oktarina, H., Khairan, K., Skala, L., Mahmud, T. (2023). Endophytic bacteria as biological agents to control fusarium wilt disease and promote tomato plant growth. Physiological and Molecular Plant Pathology, 125: 101994. https://doi.org/10.1016/j.pmpp.2023.101994
[21] Wang, Z., Xu, G., Ma, P., Lin, Y., Yang, X., Cao, C. (2017). Isolation and characterization of a phosphorus-solubilizing bacterium from rhizosphere soils and its colonization of Chinese cabbage (Brassica campestris ssp. chinensis). Frontiers in Microbiology, 8: 1270. https://doi.org/10.3389/fmicb.2017.01270
[22] Karpagam, T., Nagalakshmi, P.K. (2014). Isolation and characterization of phosphate solubilizing microbes from agricultural soil. International Journal of Current Microbiology and Applied Sciences, 3(3): 601-614. https://doi.org/10.5555/20143126541
[23] Bakker, A.W., Schippers, B.O.B. (1987). Microbial cyanide production in the rhizosphere in relation to potato yield reduction and Pseudomonas SPP-mediated plant growth-stimulation. Soil Biology and Biochemistry, 19(4): 451-457. https://doi.org/10.1016/0038-0717(87)90037-x
[24] Himpsl, S.D., Mobley, H.L. (2019). Siderophore detection using chrome azurol S and cross-feeding assays. Proteus Mirabilis: Methods and Protocols, 2019: 97-108. https://doi.org/10.1007/978-1-4939-9601-8_10
[25] Glick, B.R. (2014). Bacteria with ACC deaminase can promote plant growth and help to feed the world. Microbiological Research, 169(1): 30-39. https://doi.org/10.1016/j.micres.2013.09.009
[26] Wang, C., Wang, H., Li, Y., Li, Q., Yan, W., Zhang, Y., Wu, Z., Zhou, Q. (2021). Plant growth-promoting rhizobacteria isolation from rhizosphere of submerged macrophytes and their growth-promoting effect on Vallisneria natans under high sediment organic matter load. Microbial Biotechnology, 14(2): 726-736. https://doi.org/10.1111/1751-7915.13756
[27] Putri, Z.A. (2015). Penapisan deteksi PCR ACC-deaminase dan analisis produksi auksin rizosfer tanaman padi daerah pesisir. Institut Pertanian Bogor, Bogor.
[28] Walkley, A., Black, I.A. (1934). An examination of the Degtjareff method for determining soil organic matter, and a proposed modification of the chromic acid titration method. Soil Science, 37(1): 29-38.
[29] Kjeldahl, J. (1883) A new method for the estimation of nitrogen in organic compounds. Z. Anal. Chem., 22, 366.
[30] Bremner, J.M. (1965). Total nitrogen. Methods of Soil Analysis: Part 2 Chemical and Microbiological Properties, 9: 1149-1178. https://doi.org/10.2134/agronmonogr9.2.c32
[31] Thomas, G.W. (1982). Exchangeable cations. Methods of Soil Analysis: Part 2 Chemical and Microbiological Properties, 9: 159-165. https://doi.org/10.2134/agronmonogr9.2.2ed.c9
[32] Bai, Y., Pan, B., Charles, T.C., Smith, D.L. (2002). Co-inoculation dose and root zone temperature for plant growth promoting rhizobacteria on soybean [Glycine max (L.) Merr] grown in soil-less media. Soil Biology and Biochemistry, 34(12): 1953-1957. https://doi.org/10.1016/S0038-0717(02)00173-6
[33] Sun, P., Huang, Y., Yang, X., Liao, A., Wu, J. (2023). The role of indole derivative in the growth of plants: A review. Frontiers in Plant Science, 13: 1120613. https://doi.org/10.3389/fpls.2022.1120613
[34] Zhao, Y. (2010). Auxin biosynthesis and its role in plant development. Annual Review of Plant Biology, 61(1): 49-64. https://doi.org/10.1146/annurev-arplant-042809-112308
[35] Ganesh, J., Hewitt, K., Devkota, A.R., Wilson, T., Kaundal, A. (2024). IAA-producing plant growth promoting rhizobacteria from Ceanothus velutinus enhance cutting propagation efficiency and Arabidopsis biomass. Frontiers in Plant Science, 15: 1374877. https://doi.org/10.3389/fpls.2024.1374877
[36] Zhang, B., Li, P., Wang, Y., Wang, J., Liu, X., Wang, X., Hu, X. (2021). Characterization and synthesis of indole-3-acetic acid in plant growth promoting Enterobacter sp. RSC Advances, 11: 31601-31607. https://doi.org/10.1039/d1ra05659j
[37] Zhou, X., Wu, F. (2023). Effects of microbial inoculants on agronomic characters, physicochemical properties, and nutritional qualities of lettuce and celery in hydroponic cultivation. Frontiers in Plant Science, 14: 1234567. https://doi.org/10.3389/fpls.2023.1234567
[38] Halimursyadah, H., Syamsuddin, S., Nurhayati, N., Zuliana, Z., Phonna, T. N. (2021). Interaction between type of plant growth promoting rhizobacteria and patchouli varieties on growth and yield of patchouli (Pogostemon cablin Benth.). IOP Conference Series: Earth and Environmental Science, 667(1): 012073. https://doi.org/10.1088/1755-1315/667/1/012073
[39] Erlacher, W.A., Amaral, J.F.T.D., Amaral, J.A.T.D., Christo, B.F., Ferreira, D.S., Tatagiba, S.D., Partelli, F.L. (2023). Effect of indole-3-acetic acid on growth, physiology and nutritional status of young arabica coffee plants. Coffee Science, 17: 2050. https://doi.org/10.25186/.v17i.2050
[40] Karthika, S., Midhun, S.J., Jisha, M.S. (2020). A potential antifungal and growth-promoting bacterium Bacillus sp. KTMA4 from tomato rhizosphere. Microbial Pathogenesis, 142: 104049. https://doi.org/10.1016/j.micpath.2020.104049
[41] Raio, A., Brilli, F., Neri, L., Baraldi, R., Orlando, F., Pugliesi, C., Baccelli, I. (2023). Stenotrophomonas rhizophila Ep2. 2 inhibits growth of Botrytis cinerea through the emission of volatile organic compounds, restricts leaf infection and primes defense genes. Frontiers in Plant Science, 14: 1235669. https://doi.org/10.3389/fpls.2023.1235669
[42] Sriyanti, N.L.G., Khalimi, K., Temaja, I.G.R.M. (2019). Pemanfaatan rizobakteri penghasil indole acetic acid sebagai agens penginduksi ketahanan tanaman tembakau terhadap cucumber mosaic virus (CMV). Journal of Agricultural, Food Science & Biotechnology, 8(1): 54620.
[43] Halimursyadah, H., Hafsah, S., Nasution, S.N., Zuraida, Z. (2022). Pengaruh rizobakteri indigenous terhadap serangan penyakit budok, enzim peroksidase, dan pertumbuhan setek nilam Aceh. Kultivasi, 21(1): 81-87.
[44] Kumar, R., Singh, A., Shukla, E., Singh, P., Khan, A., Singh, N.K., Srivastava, A. (2024). Siderophore of plant growth promoting rhizobacterium origin reduces reactive oxygen species mediated injury in Solanum spp. caused by fungal pathogens. Journal of Applied Microbiology, 135(2): lxae036. https://doi.org/10.1093/jambio/lxae036
[45] Hofmann, M., Heine, T., Malik, L., Hofmann, S., Joffroy, K., Senges, C.H.R., Tischler, D. (2021). Screening for microbial metal-chelating siderophores for the removal of metal ions from solutions. Microorganisms, 9(1): 111. https://doi.org/10.3390/microorganisms9010111
[46] Kesaulya, H., Hasinu, J.V., Tuhumury, G.N. (2018). Potential of Bacillus SPP produces siderophores insuppressing thewilt disease of banana plants. IOP Conference Series: Earth and Environmental Science, Semarang, Indonesia, 102: 012016. https://doi.org/10.1088/1755-1315/102/1/012016
[47] Zhang, W., Zhang, Y., Wang, X., Ding, F., Fu, Y., Zhao, J., Chen, X. (2017). Siderophores in clinical isolates of Klebsiella pneumoniae promote ciprofloxacin resistance by inhibiting the oxidative stress. Biochemical and Biophysical Research Communications, 491(3): 855-861. https://doi.org/10.1016/j.bbrc.2017.04.108
[48] Liu, D., Yang, Q., Ge, K., Hu, X., Qi, G., Du, B., Ding, Y. (2017). Promotion of iron nutrition and growth on peanut by Paenibacillus illinoisensis and Bacillus sp. strains in calcareous soil. Brazilian Journal of Microbiology, 48(4): 656-670. https://doi.org/10.1016/j.bjm.2017.02.006
[49] Baune, M., Qi, Y., Scholz, K., Volmer, D.A., Hayen, H. (2017). Structural characterization of pyoverdines produced by Pseudomonas putida KT2440 and Pseudomonas taiwanensis VLB120. BioMetals, 30: 589-597. https://doi.org/10.1007/s10534-017-0029-7
[50] Gáll, T., Lehoczki, G., Gyémánt, G., Emri, T., Szigeti, Z. M., Balla, G., Pócsi, I. (2016). Optimization of desferrioxamine E production by Streptomyces parvulus. Acta Microbiologica et Immunologica Hungarica, 63(4): 475-489. https://doi.org/10.1556/030.63.2016.029
[51] Sudewi, S., Patandjengi, B., Ala, A., Bdr, M. F., Saleh, A.R., Ratnawati, R. (2022). Siderophore production of the rhizobacteria isolated from local “Kamba” rice plants, Poso regency in central Sulawesi. Agric, 34(2): 225-238. https://doi.org/10.24246/agric.2022.v34.i2.p225-238
[52] Huo, Y., Kang, J.P., Ahn, J.C., Kim, Y.J., Piao, C.H., Yang, D.U., Yang, D.C. (2021). Siderophore-producing rhizobacteria reduce heavy metal-induced oxidative stress in Panax ginseng Meyer. Journal of Ginseng Research, 45(2): 218-227. https://doi.org/10.1016/j.jgr.2019.12.008
[53] Maulina, N.M.I., Suprapta, D.N., Temaja, I.G.R.M., Adnyana, I.M., Suriani, N.L. (2022). Rhizobacteria of Bali with obvious growth-promoting properties on corn (Zea mays L.). Frontiers in Sustainable Food Systems, 6: 899736. https://doi.org/10.3389/fsufs.2022.899736
[54] Tian, J., Ge, F., Zhang, D., Deng, S., Liu, X. (2021). Roles of phosphate solubilizing microorganisms from managing soil phosphorus deficiency to mediating biogeochemical P cycle. Biology (Basel), 10(2): 158. https://doi.org/10.3390/biology10020158
[55] Pande, A., Pandey, P., Mehra, S., Singh, M., Kaushik, S. (2017). Phenotypic and genotypic characterization of phosphate solubilizing bacteria and their efficiency on the growth of maize. Journal of Genetic Engineering and Biotechnology, 15: 379-391. https://doi.org/10.1016/j.jgeb.2017.06.005
[56] Rawat, P., Shankhdhar, D., Shankhdhar, S.C. (2020). Plant growth-promoting rhizobacteria: A booster for ameliorating soil health and agriculture production. Soil Health, 59: 47-68. https://doi.org/10.1007/978-3-030-44364-1_3
[57] Sharma, L.K., Sharma, A.K., Ogram, A., Singh, H. (2023). Improving biological nitrogen fixation to improve soil nutrient status. EDIS, 2023(4): 71. https://doi.org/10.32473/edis-ss714-2023
[58] Kuan, K.B., Othman, R., Abdul Rahim, K., Shamsuddin, Z.H. (2016). Plant growth-promoting rhizobacteria inoculation to enhance vegetative growth, nitrogen fixation and nitrogen remobilisation of maize under greenhouse conditions. PloS One, 11(3): e0152478. https://doi.org/10.1371/journal.pone.0152478
[59] Threatt, S., Rees, D. (2022). Biological nitrogen fixation in theory, practice, and reality: A perspective on the molybdenum nitrogenase system. FEBS Letters, 597: 45-58. https://doi.org/10.1002/1873-3468.14534
[60] Nievas, F., Revale, S., Foresto, E., Cossovich, S., Puente, M., Alzari, P., Bogino, P. (2022). Complete genome sequence of Bradyrhizobium sp. strain C-145, a nitrogen-fixing rhizobacterium used as a peanut inoculant in Argentina. Microbiology Resource Announcements, 11(8): e00505-22. https://doi.org/10.1128/mra.00505-22
[61] Bandeppa, S., Phule, A.S., Barbadikar, K.M., Govindannagari, R., Kavuru, V.P.B., Mandal, P.K., Chandran, L.P. (2022). Draft genome sequence of Paenibacillus sonchi Iirrbnf1, a nitrogen-fixing and plant growth-promoting bacterium isolated from rice Rhizosphere. Microbiology Resource Announcements, 11(5): e00126-22. https://doi.org/10.1128/mra.00126-22
[62] Kuo, W.H., Small, L.L., Olsen, K.M. (2023). Variable expression of cyanide detoxification and tolerance genes in cyanogenic and acyanogenic white clover (Trifolium repens). American Journal of Botany, 110(10): e16233. https://doi.org/10.1002/ajb2.16233
[63] Mekonnen, H., Kibret, M., Assefa, F. (2022). Plant growth promoting rhizobacteria for biocontrol of tomato bacterial wilt caused by Ralstonia solanacearum. International Journal of Agronomy, 2022(1): 1489637. https://doi.org/10.1155/2022/1489637
[64] Khalil, M.S.M., Hassan, M.H.A.R., Mahmoud, A.F., Morsy, K.M.M. (2022). Involvement of secondary metabolites and extracellular lytic enzymes produced by plant growth promoting rhizobacteria in inhibiting the soilborne pathogens in Faba Bean Plants. Jurnal Hama Dan Penyakit Tumbuhan Tropika, 22(2): 100-108. https://doi.org/10.23960/jhptt.222100-108
[65] Halimursyadah, H., Syafruddin, S., Syamsuddin, S., Sriwati, R. (2023). Screening of indigenous rhizobacteria isolates from patchouli rhizosphere producing HCN, siderophores and chitinolytic enzymes. IOP Conference Series: Earth and Environmental Science, 1183: 012096. https://doi.org/10.1088/1755-1315/1183/1/012096
[66] Safriani, S., Fitri, L., Ismail, Y. (2020). Exploration of IAA and HCN Producing Rhizobacteria from Cassava (Manihot esculenta Crantz) Rhizosphere. Jurnal Biologi Indonesia, 16: 71-78.
[67] Shahid, M., Singh, U.B., Khan, M.S., Singh, P., Kumar, R., Singh, R.N., Singh, H.V. (2023). Bacterial ACC deaminase: Insights into enzymology, biochemistry, genetics, and potential role in amelioration of environmental stress in crop plants. Frontiers in Microbiology, 14: 1132770. https://doi.org/10.3389/fmicb.2023.1132770
[68] Yoolong, S., Kruasuwan, W., Thanh Phạm, H.T., Jaemsaeng, R., Jantasuriyarat, C., Thamchaipenet, A. (2019). Modulation of salt tolerance in Thai jasmine rice (Oryza sativa L. cv. KDML105) by Streptomyces venezuelae ATCC 10712 expressing ACC deaminase. Scientific Reports, 9(1): 1275. https://doi.org/10.1038/s41598-018-37987-5
[69] Sagar, A., Rathore, P., Ramteke, P.W., Ramakrishna, W., Reddy, M.S., Pecoraro, L. (2021). Plant growth promoting rhizobacteria, arbuscular mycorrhizal fungi and their synergistic interactions to counteract the negative effects of saline soil on agriculture: Key macromolecules and mechanisms. Microorganisms, 9(7): 1491. https://doi.org/10.3390/microorganisms9071491
[70] Danish, S., Zafar-Ul-Hye, M., Mohsin, F., Hussain, M. (2020). ACC-deaminase producing plant growth-promoting rhizobacteria and biochar mitigate adverse effects of drought stress on maize growth. PLoS One, 17(11): e0277591. https://doi.org/10.1371/journal.pone.0277591
[71] Heydarian, Z., Yu, M., Gruber, M., Glick, B.R., Zhou, R., Hegedus, D.D. (2016). Inoculation of soil with plant growth promoting bacteria producing 1-aminocyclopropane-1-carboxylate deaminase or expression of the corresponding acdS gene in transgenic plants increases salinity tolerance in Camelina sativa. Frontiers in Microbiology, 7: 1966. https://doi.org/10.3389/fmicb.2016.01966
[72] Maulidia, V. (2022). Seed treatment using rhizobacteria as plant growth promotion of two chili variety (Capsicum annuum L.). IOP Conference Series: Earth and Environmental Science, 012060. https://doi.org/10.1088/1755-1315/951/1/012060
[73] Gupta, R., Noureldeen, A., Darwish, H. (2021). Rhizosphere mediated growth enhancement using phosphate solubilizing rhizobacteria and their tri-calcium phosphate solubilization activity under pot culture assays in Rice (Oryza sativa.). Saudi Journal of Biological Sciences, 28(7): 3692-3700. https://doi.org/10.1016/j.sjbs.2021.05.052
[74] Han, J., Sun, L., Dong, X., Cai, Z., Sun, X., Yang, H., Song, W. (2005). Characterization of a novel plant growth-promoting bacteria strain Delftia tsuruhatensis HR4 both as a diazotroph and a potential biocontrol agent against various plant pathogens. Systematic and Applied Microbiology, 28(1): 66-76. https://doi.org/10.1016/j.syapm.2004.09.003
[75] Hou, Q., Wang, C., Guo, H., Xia, Z., Ye, J., Liu, K., Ding, Y. (2015). Draft genome sequence of Delftia tsuruhatensis MTQ3, a strain of plant growth-promoting rhizobacterium with antimicrobial activity. Genome Announcements, 3(4): 1128. https://doi.org/10.1128/genomea.00822-15
[76] Guo, H., Yang, Y., Liu, K., Xu, W., Gao, J., Duan, H., Wang, C. (2016). Comparative genomic analysis of Delftia tsuruhatensis MTQ3 and the identification of functional NRPS genes for siderophore production. BioMed Research International, 2016(1): 3687619. https://doi.org/10.1155/2016/3687619
[77] Ghadamgahi, F., Tarighi, S., Taheri, P., Saripella, G.V., Anzalone, A., Kalyandurg, P.B., Vetukuri, R.R. (2022). Plant growth-promoting activity of Pseudomonas aeruginosa FG106 and its ability to act as a biocontrol agent against potato, tomato and taro pathogens. Biology, 11(1): 140. https://doi.org/10.3390/biology11010140
[78] Laid, B., Kamel, K., Mouloud, G., Manel, S., Walid, S., Amar, B., Faiçal, B. (2016). Effects of plant growth promoting rhizobacteria (PGPR) on in vitro bread wheat (Triticum aestivum L.) growth parameters and biological control mechanisms. Advances in Microbiology, 6(9): 677-690. https://doi.org/10.4236/aim.2016.69067
[79] Azizah, N., Kurniawan, T., Halimursyadah, H. (2022). Kemampuan isolat rizobakteri indigenous sebagai agen biofertilizer terhadap pertumbuhan beberapa varietas nilam Aceh (The ability of indigenous rhizobacteria isolates as biofertilizer agents on growth of some Aceh patchouli varieties). Jurnal Ilmiah Mahasiswa Pertanian, 7(4): 151-160.
[80] Agbodjato, N.A., Babalola, O.O. (2024). Promoting sustainable agriculture by exploiting plant growth-promoting rhizobacteria (PGPR) to improve maize and cowpea crops. PeerJ, 12: e16836. https://doi.org/10.7717/peerj.16836
[81] Compant, S., Samad, A., Faist, H., Sessitsch, A. (2019). A review on the plant microbiome: Ecology, functions, and emerging trends in microbial application. Journal of Advanced Research, 19: 29-37. https://doi.org/10.1016/j.jare.2019.03.004
[82] Berendsen, R.L., Pieterse, C.M., Bakker, P.A. (2012). The rhizosphere microbiome and plant health. Trends in Plant Science, 17(8): 478-486.