Darwin*![]() | Ramayanty Bulan
| Ramayanty Bulan![]() | Salwa Rana Fitria
| Salwa Rana Fitria![]() | Andriy Anta Kacaribu
| Andriy Anta Kacaribu![]()
© 2025 The authors. This article is published by IIETA and is licensed under the CC BY 4.0 license (http://creativecommons.org/licenses/by/4.0/).
OPEN ACCESS
The main purpose of this experimental study was to assess the accumulation of organic acids and their mitigation in rumen cultures fermenting silages. The silages used in the rumen fermentation were composed of corn stalks, bagasse, and rice straw. The rumen fermentation was conducted in vitro using batch systems at 38.5 ± 0.5℃ for 48 hours. The results showed that within 4 hours of incubation, the rumen culture fermenting bagasse silage produced 40% more VFA than other silages. Lactic acid build-up occurred exclusively in the rumen culture fermenting bagasse silage. Lactic acid production peaked after 8 hours of incubation, reaching a pH of 5.5. The addition of bentonite to the rumen culture fermenting bagasse silage reduced lactic acid concentration by 65%. Supplementing the feed with 10% bentonite can significantly increase pH from 5.5 to 6.2, potentially mitigating ruminal acidosis.
silages, feedstuffs, acidosis, organic acid build-up
Fermentation is an essential process in the rumen that degrades and converts feedstuffs into energy sources utilized by the host [1]. The fermentation characteristics of complex feeds can be studied in vivo and in vitro [2]. As in vivo assessment of rumen fermentation characteristics is expensive, labor-intensive, and difficult to standardize, in vitro studies have been established to evaluate the fermentation process in the rumen [3]. In vitro fermentation is an important technique to assess the nutritional value of complex feeds or specific feedstuffs and to observe fermentation processes in the rumen. Furthermore, in vitro rumen fermentation can be used to evaluate nutrient metabolism mechanisms and investigate shifts in metabolic pathways and metabolite formation that may affect the host’s growth, reproduction, and health [4].
Ruminants are normally adapted to consume and digest forage feeds [5]. However, in modern animal production systems and intensive fattening systems, ruminants are typically fed with high-grain diets and processed feedstuffs [6, 7]. This practice is applied to enhance the energy consumption of the ruminants and maximize their productivity [8]. Prolonged excessive feeding of non-forage or low-fiber diets, such as high-grain and processed feeds, may lead to the accumulation of organic acids in the rumen [9, 10]. An accumulation of organic acids may cause a drop in pH in the rumen culture leading to a metabolic disorder called acidosis [11]. Rumen acidosis or digestive disorder frequently occurs when the rumen is continuously supplied with easily digestible carbohydrates mainly cereal grains [12, 13].
Cereal grains or starchy feedstuffs supplied to the rumen are easily degraded and rapidly converted into organic acids, mainly lactic acid [14], where the rate of lactic acid formation frequently surpasses the rumen’s absorption capacity [15]. This acid build-up may decrease ruminal pH and generate an acidic state in the rumen culture. Hence, this condition would create some side effects for the host including changes in the water balance, food intake, and digestibility, and shifts of the acid-base balance in rumen fluid [7, 16]. Further, rumen acidosis certainly would affect the performance and productivity of the ruminants [17]. Acute acidosis may cause morbidity and mortality in ruminant populations [18]. Hence, feeding of readily fermentable materials particularly carbohydrate-based feedstuffs should be managed properly to prevent the risk of organic acids accumulating in the rumen culture. Also, feasible buffers or buffer agents should be considered to mitigate rumen acidosis [19]. In addition to cereal grain feeds, silage is an alternative feed used by cattle growers to increase their productivity [20, 21]. Currently, forages fed as silage have remained popular for modern dairy farms as they could reduce the loss of nutrients from harvest via storage, enable easier handling and feeding, are more efficient, retain more nutrients, and have higher nutritional value than dry forages [20]. Silage is a fermented green feedstuff for ruminants, typically made from green foliage crops, grasses, or other lignocellulosic materials, such as corn stover, wheat straw, and rice straw [22]. Ensilage or ensiling is a preserving process of the green grass crops and lignocellulosic materials into silo or fermentation chambers under anaerobic conditions [23]. The production of silage typically occurs under the acidification state. At this stage, the pH of the silage is somewhat low (pH < 5.0) due to the formation of organic acids primary lactic acid, and acetic acid [24]. A rapid decrease in silage pH may assist in restricting the degradation of protein in the fermentation chamber via inactivating plant proteases [25]. As the produced silage is typically quite acidic [26], prolonged feeding of this fodder to the ruminants should be monitored properly and evaluated to avoid any potential rumen acidosis. The present study aims to assess the risk of ruminal acidosis in rumen cultures fermenting various types of silage. A potential acidosis mitigation technique will also be investigated in this study, in which a feasible buffer material, such as bentonite, is introduced to the rumen cultures fermenting silages. Digestion efficiency will also be assessed to evaluate feed digestibility during the acidification process.
2.1 Rumen fluid collection and preparation
Rumen fluid used for the in-vitro study of rumen culture fermenting various types of silages was taken from cattle slaughtered in a commercial slaughterhouse (with undefined diet) The slaughterhouse is located in Peunayong, Banda Aceh City, Aceh Province, Indonesia. Soon after the collection, the rumen fluid was stored in the bottle and kept under the warm temperature of 38.5 ± 0.5℃ before the start of the series of experiments to maintain microbial activity [27-29]. The collected rumen fluid was screened for removing any particles before use in the experiments.
2.2 Substrates preparation
Some substrates utilized for the in-vitro rumen fermentation are various kinds of silages made from lignocellulosic biomass including corn stover and stalks, bagasse, and rice straw. Before the ensiling process, these materials were reduced and chopped into 3 cm. As the feedstock possesses somewhat low sugar content, each type of lignocellulosic material (1 kg) was then added with molasses (100 ml) to enhance the rate of the fermentation process. The mixture was then inoculated with 10% of the effective microorganisms (EM4). The mixture was preserved for 21 incubation days for generating silages [30].
2.3 Design and procedures of the experiments
2.3.1 Experimental setup
The experiments were carried out in batch trials with various types of silage generated and bentonite supplementation. The batch tests were performed utilizing a thermostatic water bath at 38.5 ± 0.5℃, close to the rumen temperature between 37.8 and 40℃ [31]. The in-vitro rumen fermentation lasted 48 hours of incubation allowing rumen microorganisms to acclimatize to the anoxic state and providing adequate time for the microbes to degrade and convert all substrate into fermentation end-products. In these trials, fermentation was conducted in batch digesters, and the working volume established was 0.1 liter. The substrate concentration introduced was 100 g/l. Each fermentation chamber was fed 10 g of substrate in 100 ml of the filtered rumen fluid. The substrate was varied with different types of silages obtained to evaluate their fermentation characteristics.
2.3.2 Evaluation of various substrate concentrations
In the study of rumen culture fermenting different types of silages, some tests introduced were a blank digester consisting solely of rumen fluid (S0) as a control, rumen culture fermenting silage of corn stalk (S1), rumen culture fermenting silage of bagasse (S2), and rumen culture fermenting silage of rice straw (S3). This trial aims to evaluate the rumen fermentation characteristics of each type of silage made from the lignocellulosic biomass. Besides, this test assessed any potential acidosis caused by feeding silage to the rumen culture. No acid and/or alkaline solutions were introduced to the culture.
To mitigate any potential acidosis in the rumen culture fermenting silages, bentonite was supplemented to the culture as a buffer. Each silage (8 g) and bentonite (2 g) were mixed into the rumen fluid (100 ml), and thereby the total concentration was 100 g/l. The design of experiments included the rumen culture fermenting corn stalk silage supplemented with bentonite (S1B), rumen culture fermenting bagasse silage added with bentonite (S2B), and rumen culture fermenting silage of rice straw supplemented with bentonite (S3B). A control test was also introduced in which the rumen culture was added with mere bentonite (S0B). The bentonite used in this experiment was a commercial calcium bentonite clay for animal feed. During the fermentation, the pH of the culture was not maintained at any particular level. Hence, no other chemicals were supplemented into the rumen culture.
2.4 Analytical methods
Each sample collected from the experiments including samples before and after in-vitro rumen fermentation was periodically measured to evaluate their pH level as the main indicator of rumen acidosis. The pH measurement was carried out using a Laboratory Benchtop pH Meter Multifunction Complete Probe, Milwaukee PRO, and to ensure accurate and precise measurement the pH meter was regularly calibrated with pH buffer solutions of 4.0, 7.0, and 10 [32, 33]. Alkalinity was analyzed to assess the buffering capacity of rumen fluid and substrates used to neutralize acids and bases formed during the in-vitro rumen fermentation of silages. The total alkalinity of each sample was measured by titration method using the pH meter and hydrochloric acid reagent [34, 35].
Samples of the collected rumen fluid and rumen culture fermenting silages were analyzed for their ammonia level utilizing NH3/NH4+ test kit and reagent according to the colorimetric procedure [36, 37]. The determination of protein content (crude) of the substrates used was carried out based on the block digestion method according to the AOAC official method 2001.11 [38, 39]. The substrate's fiber content (crude) was analyzed according to the AOAC official method 962.09 [40].
Each sample collected was dried for 24 hours under the temperature of 105 ± 0.5℃ utilizing a laboratory drying oven to measure their moisture content (MC), dry matter (DM) or total solid [41, 42]. The dried samples were then placed in the muffle furnace for 12-18 hours at around 575 ± 25℃ for ash content determination. All solid analysis was carried out according to the standard methods of APHA [43]. Digestion efficiency analysis was performed to evaluate the effectiveness of substrate degradation during in-vitro rumen fermentation [44].
Organic acids including volatile fatty acids (VFA) and lactic acid were analyzed from the fermented samples to evaluate the types of organic acids generated and build-up during the fermentation process of silages. The analysis was performed using a titrimetric method. The solution of NaOH (0.1 M) was utilized as a titrant and filled into the burette tube. Before the measurement, some drops of phenolphthalein indicator were added to the samples analyzed [45, 46]. The measurement of lactic acid formation and soluble carbohydrates of the fermented samples was also validated using a portable sensitive Lactate, Gluco-Biosensor [47, 48]. To ensure the data generated was reproducible, all samples measured were replicated.
The current study uses batch tests to evaluate the in-vitro rumen fermentation of various types of silage made of lignocellulosic biomass. The results showed that the pH of the rumen fluid used for the experiments was about 7.41 (Table 1), which was slightly higher than the optimal rumen pH between 6.4 and 7.0 [49, 50]. The pH level of the rumen fluid used in this study suggested that no organic acid build-up and/or ruminal acidosis occurred previously in the ruminant, and thereby would be feasibly used for assessing the impact of various types of silage feed in the rumen fermentation in-vitro.
Table 1. Physicochemical properties of rumen fluid and substrates used for rumen fermentation in-vitro
|
Parameters |
Unit |
Rumen Fluid |
Corn Stalk Silage |
Bagasse Silage |
Rice Straw Silage |
|
Dry matter |
% |
2.89 |
45.3 |
61.1 |
64.9 |
|
Moisture content |
% |
97.11 |
54.7 |
38.9 |
35.1 |
|
pH |
- |
7.41 |
3.89 |
3.91 |
4.52 |
|
Alkalinity |
mg/l |
1.4 |
3.0 |
2.0 |
3.0 |
|
Protein (crude) |
% |
0.43 |
3.51 |
3.3 |
4.31 |
|
Fiber (crude) |
% |
4.98 |
15.39 |
14.72 |
13.79 |
|
Ash content |
% |
0.51 |
6.89 |
5.01 |
14.80 |
|
Ammonia |
mg/l |
60 |
- |
- |
- |
|
VFA |
g/l |
6.38 |
- |
- |
- |
As presented in Table 1, the ammonia content of the rumen fluid used in in-vitro rumen fermentation was around 60 mg/l. Some studies revealed that the minimum ammonia concentration in the rumen should be between 40 and 240 mg/l to support the microbes in the rumen to grow well and active [51, 52]. This suggested that the ammonia level in the rumen fluid used in this study was still in the minimum acceptable concentration range for supporting the growth of ruminal microbial consortia, which was feasible for performing rumen fermentation in vitro.
Table 1 shows that silages used as substrates for rumen fermentation in vitro possess significant amounts of valuable nutrients, including protein (3.3-3.51%) and fiber (13.80-15.40%), minerals represented in ash content (5.01-14.80%), and dry matter (45.3-64.9%). These nutrients benefit the hosts, enhancing microbial activity and improving substrate digestion in the rumen ecosystem [53]. Besides, the study mentioned that supplementation of fermented protein products may enhance digestibility, reduce antinutritional factor (ANF), and increase antioxidant and other biological activities, such as probiotics, and prebiotics [54].
The results show that the pH of the silages was somewhat acidic between 3.89 and 4.52. A study mentioned that good-quality silages would be associated with low pH [25]. The study added that the silages should have a pH of 3.8 to 4.2. This study shows that the nutritional value of silages may be associated with the pH level. As depicted in Table 1, the higher the pH, the lower the fiber content of the silages obtained. In this study, corn stalk silage has a lower pH (3.89) than other silages but its fiber content is higher (15.39%) than the others. This study also shows that silages with low pH have lower protein content than silages with high pH. In this study, rice straw silage with pH 4.52 has a higher protein content (4.31%) than other silages with lower pH content. This indicated that low pH may restrict the synthesis of protein.
The current study revealed that the VFA concentration of the rumen fluid used for the rumen fermentation in vitro was around 6.38 g/l. The study mentioned that the VFA concentration of the rumen was normally in the range of 5-10 g/l [55]. This suggested that the rumen fluid collected was feasibly used for studying in-vitro rumen fermentation of silages. To maintain balance in the rumen ecosystem and support the optimal growth and activities of the rumen microorganisms, the VFA of the rumen culture should be kept within those normal ranges. This is because when the VFA concentration of the rumen is higher than that of the normal ranges, the pH of the rumen may decrease and thereby potentially induce rumen acidosis [55, 56].
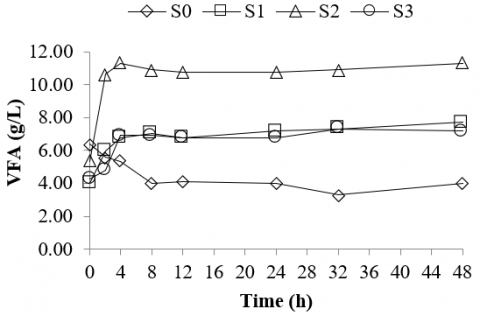
The results reveal that the rumen culture fermenting silage of bagasse produced higher VFA than that of the rumen culture fermenting silage of corn stalk and rice straw (Figure 1). Interestingly, the VFA build-up in the rumen culture fermenting bagasse's silage occurred within 2 hours of incubation. As depicted in Figure 1, the VFA concentration produced in the rumen culture fermenting silage of bagasse was two times higher (11 g/l) than that of the rumen culture fermenting silage of corn stalk and rice straw (6.8 – 6.9 g/l) and the control or rumen culture only (5.3 g/l). Higher VFA concentration in the fermentation culture frequently led to pH depression [57, 58], and potentially generated acidosis [56]. Hence, supplementing a suitable buffer material, such as bentonite would potentially enhance the buffer capacity and prevent pH depression in the VFA-producing culture.
Figure 1. VFA of rumen culture fermenting various types of silages
Solely of rumen fluid (S0) as a control, rumen culture fermenting silage of corn stalk (S1), rumen culture fermenting silage of bagasse (S2), and rumen culture fermenting silage of rice straw (S3).
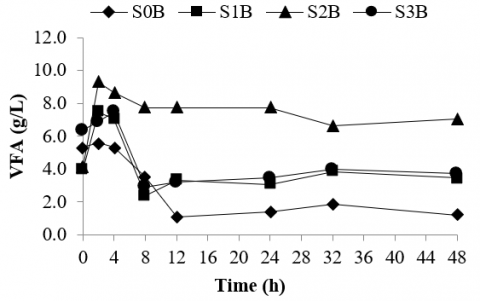
The results show that after 8 hours of incubation, the concentration of VFA in all sample tests tended to decrease (Figure 2). As depicted in Figure 2, from 2 to 12 hours of the fermentation period, VFA in the rumen culture fermenting silage of bagasse supplemented with bentonite reduced from 9.30 to 7.7 g/l while the concentration of VFA in the rumen culture fermenting silage of corn stalk and rice straw decreased from 7.0 to around 3.0 g/l and the control sample dropped from 5.6 to 1.1 g/l. This suggested that supplementation of bentonite to the rumen culture fermenting silages may reduce its VFA build-up. Even if the supplementation of bentonite did not significantly reduce the VFA formation in the rumen fermentation, the use of bentonite as a feed additive may generate some benefits to the host including enhancing buffer capacity in the rumen, improving feed efficiency and rate of gain, reduce or eliminate the toxicity [59]. The study reported that supplementing bentonite between 0.2 and 10% of the feeds is still safe for the ruminants [60]. This agrees with the current study that supplemented merely 2% of bentonite to the silage feeds for mitigating and eradicating potential ruminal acidosis.
Figure 2. VFA of rumen culture fermenting various types of silages supplemented with bentonite
The rumen culture was added with mere bentonite (S0B) as a control, the rumen culture fermenting corn stalk silage supplemented with bentonite (S1B), rumen culture fermenting bagasse silage added with bentonite (S2B), and rumen culture fermenting silage of rice straw supplemented with bentonite (S3B).
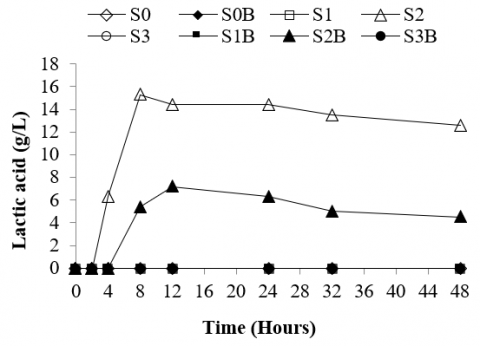
The results show that the rumen culture fermenting silage of bagasse produced a significant amount of lactic acid. However, no lactic acid production was found in the rumen culture fermenting silages made of corn stalks and rice straw. This could be attributed that bagasse may still contain higher sugar content than other feedstocks, and thereby this would be a useful substrate for fermentative microbes to form lactic acid. Besides, within 4 hours of incubation, the rumen culture fermenting silage of bagasse produced lactic acid (6.30 g/l). The production of lactic acid in the rumen culture fermenting silage of bagasse reached a peak at 8 hours of incubation with a concentration of 15.30 g/l (Figure 3). The accumulated lactic acid frequently may generate a pH depression (Figure 4) and lead to acidosis [61]. Generating significant amounts of lactic acid in the rumen may cause ruminal acidosis [14]. Hence, lactic acid build-up in the rumen fermentation should be controlled properly to prevent severe or acute ruminal acidosis.
Supplementation of bentonite to the rumen culture fermenting silage of bagasse could reduce the formation of lactic acid in which within 8 hours of incubation lactic acid produced was merely 5.40 g/l (Figure 3). This suggested that supplementing bentonite to the silage feed fermented in the rumen may significantly reduce the lactic acid production and prevent its accumulation during the fermentation. Hence, potential ruminal acidosis in the rumen culture fermenting silage of bagasse could be mitigated by supplementing 10% of bentonite to the feed. This is crucial since feeding silage made of bagasse to the ruminants would be somewhat risky due to the potential lactic acid accumulated in the rumen. This is because supplementation of bentonite may enhance buffer capacity in the rumen, and thereby the acids generated during the rumen fermentation could be neutralized. This study agrees with the study reporting that bentonite supplemented with the ruminants fed with high-concentrate diets may have positive effects, such as maintaining optimal pH and preventing acidosis [62].
Figure 3. Lactic acid production of rumen culture fermenting various types of silages supplemented with bentonite
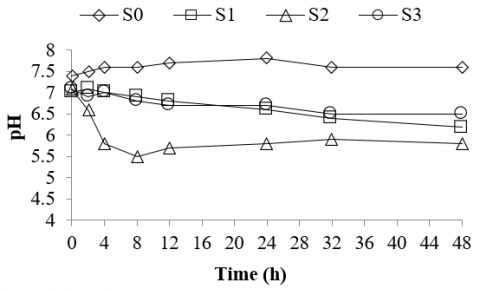
Figure 4. pH of rumen culture fermenting various types of silages
The current study results show that any potential acidosis may occur when the rumen culture is fed with silage particularly the silage made of bagasse. In this study, the pH of the rumen culture fermenting silage made of bagasse dropped significantly from 7.2 to 5.5 within 8 hours of incubation while the pH of the control was stable between 7.5 and 7.6 (Figure 4). This indicated that ruminal acidosis may occur within less than 10 hours of incubation when the rumen was fed with silage made of bagasse. This is somewhat different from the rumen culture fermenting silages made of corn stalk and rice straw in which within 8 hours of incubation, the pH of the culture was still stable between 7.0 and 6.8. This may be attributed to the characteristics of the silages made of corn stalk and rice straw that have higher alkalinity (3.0 mg/l) than that of the silages made of bagasse (2.0 mg/l). A higher alkalinity of the substrate fed would be preferable to maintain a fairly constant pH in the fermentation culture. This is because the substrates or cultures with high alkalinity typically may be able to neutralize stronger acids by accepting proton or H+ ions released during the fermentation processes [63].
The results reveal that the rumen culture fermenting silage made of bagasse does not have sufficient capability to neutralize acids formed during the fermentation. As a consequence, within 48 hours of the incubation period, the pH of the rumen culture fermenting silage of bagasse cannot recover, but it remains stable between 5.5 and 5.8. When the pH in the rumen remains low, a transient imbalance between organic acid production and absorption may occur and lead to rumen metabolic disorder or acidosis [56]. The low pH could be linked to the organic acid build-up during the rumen fermentation in which an acidic state due to organic acids (i.e. VFA and lactic acid) accumulated in the rumen culture would significantly depress its pH level [14]. The study reported that the production of lactic acid would be optimal when the pH of the culture was between 5.5 and 5.8 [64, 65]. At this stage, the rumen culture may be upset and lead to metabolic disorder or rumen acidosis. This agrees with the study reporting that a disorder of ruminal fermentation or sub-acute ruminal acidosis (SARA) occurs when the pH of the rumen remains in the range of 5.2 and 6.0 for a prolonged period [66, 67].
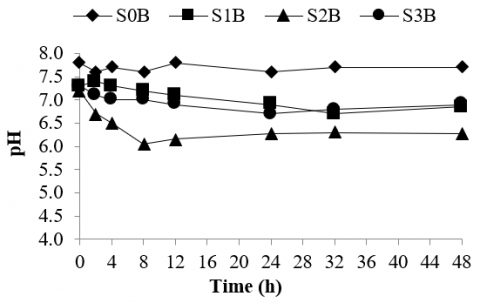
Figure 5. pH of rumen culture fermenting various types of silages supplemented with bentonite
As depicted in Figure 5, within 8 hours of incubation, the pH of rumen culture fermenting silage made of bagasse decreased from 7.2 to 6.0. Then 4 hours later the pH recovered and increased from 6.0 to 6.2. Hence, supplementation of bentonite may relieve and mitigate sub-acute ruminal acidosis in the rumen culture fermenting silage of bagasse. Besides, during the fermentation process, the pH of the rumen culture fermenting silages made of corn stalk and rice straw supplemented with bentonite remained stable between 6.80 and 7.0 with no significant drop in pH throughout the process. Meanwhile, the pH of the control was somewhat alkaline and stable in the range between 7.6 and 7.7. This suggested that supplementation of bentonite to the rumen culture fermenting silages may significantly impact the risk of ruminal acidosis by enhancing the buffering capacity of the culture and improving the pH level. Hence, this practice may generate a synergetic effect on the rumen fermentation culture but potentially prevent a significant decrease in the pH, metabolic disorder, and rumen acidosis [68].
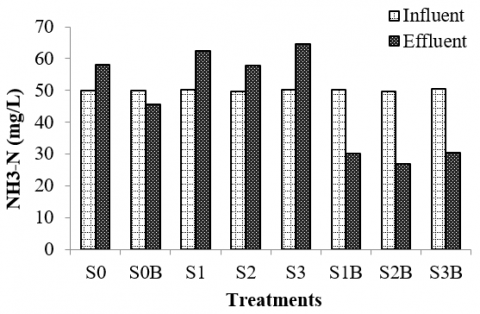
The study mentioned that the minimum concentration of ammonia in rumen fluid for rumen microorganisms to grow well is around 50 mg/l [55]. This agrees with the current study, in which the ammonia in the influent was between 49 and 50 mg/l. The current study results show that fermentation may alter ammonia concentration in the rumen culture. As depicted in Figure 6, after 48 hours of fermentation period ammonia concentration in the rumen culture fermenting silages increased from 50 mg/l to around 60 mg/l in which the rumen culture fermenting silage of rice straw had a higher level of ammonia (65 mg/l) compared to the others. This could be related to the level of pH in the rumen culture fermenting silage of rice straw (pH 6.5) which was higher than that of the rumen culture fermenting silage of corn stalk (pH 6.2) and bagasse (pH 5.8). On the other hand, the concentration of ammonia in the rumen culture fermenting silages supplemented with bentonite decreased significantly from 50 mg/l to around 30 mg/l. Besides, the control reactor also depicts a reduction of ammonia when the rumen culture was supplemented with bentonite from 60 to 45 mg/l.This would be attributed to the characteristic of bentonite, which has a high ammonia adsorption capacity and can decrease ammonium nitrogen in the culture [69].
Figure 6. Ammonia of rumen culture fermenting various types of silages supplemented with bentonite
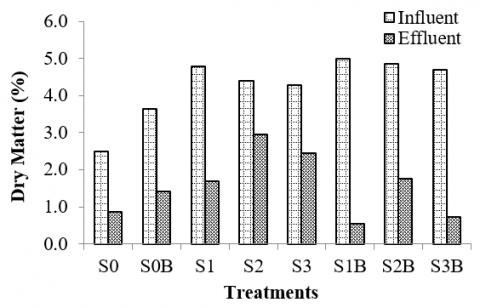
Figure 7. Profile of dry matter of rumen culture fermenting various types of silages supplemented with bentonite
The results show that rumen fermentation in-vitro successfully reduced the dry matter of the rumen culture fed with various types of silages and supplemented with bentonite. As depicted in Figure 7, after 48 hours of incubation the rumen culture fermenting corn stalk silage had lower dry matter (1.7% DM) compared to that of the rumen culture fermenting silages of bagasse (3.0% DM) and rice straw (2.4% DM). The supplementation of bentonite to the rumen culture fermenting corn stalk silage had significantly improved the digestion of corn stalk silage in which its dry matter decreased considerably from 5.0% to 0.5%. This suggested that the corn stalk silage was the substrate that was highly degraded as well as digested during the rumen fermentation. This indicates that corn stalk silage is a potential feed that can be used to substitute corn silage. This is because corn stalk instead of corn grain for producing silages would be significant as corn stalk typically is discarded during harvesting corn. A study mentioned that replacing corn silage with corn stalk silage in the finishing feed had no effect or negative impacts on the proximate nutrients, fatty acids profile, pH, and water-holding capacity [70].
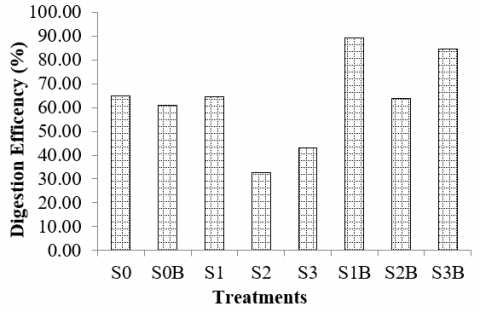
The results of the current study reveal that supplementation of bentonite may enhance the digestion in the rumen culture fermenting silages. As shown in Figure 8, when the rumen culture fermenting bagasse silage was supplemented with bentonite, its digestion efficiency increased from 33 to 64%. This is because bentonite supplementation may improve the buffer capacity of the rumen culture fermenting silages. In this study, the pH of the rumen culture fermenting silage of bagasse increased significantly from 5.8 to 6.28 after the supplementation of bentonite (Figures 4 and 5). The results agree with the studies reporting that the primary fibrolytic bacteria are intolerant to the low pH of the rumen, and rumen fiber digestion may significantly decrease when the pH of the rumen is too low or below critical levels [71]. Hence, by combining silages and bentonite, ruminal pH would be improved and support rumen microbes in digesting and degrading the substrates.
Figure 8. Digestion efficiency of various substrates fermented in the rumen culture
These experimental results indicate that ruminal acidosis may occur in the rumen fermentation of silages made of lignocellulosic materials, particularly bagasse. Rumen fermentation of bagasse silage generates organic acid build-up including VFA and lactic acid. Lactic acid was generated in the rumen culture fermenting bagasse silage within merely 4 hours of incubation (6.3 g/l) and was accumulated after 8 hours of fermentation period (15.3 g/l). The supplementation of bentonite as an animal feed additive may significantly reduce lactic acid accumulation and increase pH to a safe range (6.8-7.0).
The authors acknowledge the National Research and Innovation Agency (BRIN) and the Indonesia Endowment Fund for Education (LPDP), the Republic of Indonesia for funding the research project under the scheme of Research and Innovation for Advanced Indonesia (RIIM) (Grant number of multiyear research: 82/II.7/HK/2022), The authors also thank the Institute for Research and Community Services (LPPM), Universitas Syiah Kuala.
[1] Valente, T.N.P., da Silva Lima, E., dos Santos, W.B.R., Cesário, A.S., Tavares, C.J., De Freitas, M.A.M. (2016). Ruminal microorganism consideration and protein used in the metabolism of the ruminants: A review. African Journal of Microbiology Research, 10(14): 456-464. https://doi.org/10.5897/AJMR2016.7627
[2] Cone, J.W., Van Gelder, A.H., Soliman, I.A., De Visser, H., Van Vuuren, A.M. (1999). Different techniques to study rumen fermentation characteristics of maturing grass and grass silage. Journal of Dairy Science, 82(5): 957-966. https://doi.org/10.3168/jds.S0022-0302(99)75315-4
[3] Wei, X., Ouyang, K., Long, T., Liu, Z., Li, Y., Qiu, Q. (2022). Dynamic variations in rumen fermentation characteristics and bacterial community composition during in vitro fermentation. Fermentation, 8(6): 276. https://doi.org/10.3390/fermentation8060276
[4] Joch, M., Kudrna, V., Hakl, J., Božik, M., Homolka, P., Illek, J., Tyrolová, Y., Výborná, A. (2019). In vitro and in vivo potential of a blend of essential oil compounds to improve rumen fermentation and performance of dairy cows. Animal Feed Science and Technology, 251: 176-186. https://doi.org/10.1016/j.anifeedsci.2019.03.009
[5] Mahanta, S.K., Garcia, S.C., Islam, M.R. (2020). Forage based feeding systems of dairy animals: Issues, limitations and strategies. Range Management and Agroforestry, 41(2): 188-199.
[6] Humer, E., Zebeli, Q. (2017). Grains in ruminant feeding and potentials to enhance their nutritive and health value by chemical processing. Animal Feed Science and Technology, 226: 133-151. https://doi.org/10.1016/j.anifeedsci.2017.02.005
[7] Jaramillo-López, E., Itza-Ortiz, M.F., Peraza-Mercado, G., Carrera-Chávez, J.M. (2017). Ruminal acidosis: Strategies for its control. Austral Journal of Veterinary Sciences, 49(3): 139-148. https://doi.org/10.4067/S0719-81322017000300139
[8] Molossi, L., Hoshide, A.K., Pedrosa, L.M., Oliveira, A.S.D., Abreu, D.C.D. (2020). Improve pasture or feed grain? Greenhouse gas emissions, profitability, and resource use for nelore beef cattle in Brazil’s Cerrado and Amazon Biomes. Animals, 10(8): 1386. https://doi.org/10.3390/ani10081386
[9] Millen, D.D., Pacheco, R.D.L., da Silva Cabral, L., Cursino, L.L., Watanabe, D.H.M., Rigueiro, A.L.N. (2016). Ruminal acidosis. Rumenology, 127-156.
[10] Zenker, W., Clauss, M., Huber, J., Altenbrunner-Martinek, B. (2009). Rumen pH and hoof health in two groups of captive wild ruminants. Zoo Animal Nutrition, (4): 247-254. https://doi.org/10.5167/uzh-20245
[11] Monteiro, H.F., Faciola, A.P. (2020). Ruminal acidosis, bacterial changes, and lipopolysaccharides. Journal of Animal Science, 98(8): skaa248. https://doi.org/10.1093/jas/skaa248
[12] Wu, G. (2020). Management of metabolic disorders (including metabolic diseases) in ruminant and nonruminant animals. In Animal agriculture, pp. 471-491. https://doi.org/10.1016/B978-0-12-817052-6.00027-6
[13] Darwin, Bulan, R., Humaira, T., Muliawati, A., Agustina, F., Baszanova, G.S.T. (2023). Impact of concentrate supplementation on biodegradability and acidosis in in-vitro rumen fermentation of forage. International Journal of Design & Nature and Ecodynamics, 18(4): 877-883. https://doi.org/10.18280/ijdne.180414
[14] Golder, H.M., Lean, I.J. (2024). Ruminal acidosis and its definition: A critical review. Journal of Dairy Science, 107(12): 10066-10098. https://doi.org/10.3168/jds.2024-24817
[15] Hajikolaei, M.H., Nouri, M., Afshar, F.S., Dehkordi, A.J. (2006). Effects of experimentally induced ruminal lactic acidosis on blood pH, bicarbonate and pCO2 in the sheep. Pakistan Journal of Biological Sciences, 9: 2003-2005.
[16] Faniyi, T.O., Adegbeye, M.J., Elghandour, M.M.M.Y., Pilego, A.B., Salem, A.Z.M., Olaniyi, T.A., Adediran, O., Adewumi, M.K. (2019). Role of diverse fermentative factors towards microbial community shift in ruminants. Journal of Applied Microbiology, 127(1): 2-11. https://doi.org/10.1111/jam.14212
[17] Santra, A., Karim, S.A. (2003). Rumen manipulation to improve animal productivity. Asian-Australasian Journal of Animal Sciences, 16(5): 748-763. https://doi.org/10.5713/ajas.2003.748
[18] Voulgarakis, N., Gougoulis, D., Psalla, D., Papakonstantinou, G., Angelidou-Tsifida, M., Papatsiros, V., Athanasiou, L.V., Christodoulopoulos, G. (2023). Ruminal Acidosis Part I: Clinical manifestations, epidemiology and impact of the disease. Journal of the Hellenic Veterinary Medical Society, 74(3): 5883-5891. https://doi.org/10.12681/jhvms.31237
[19] Blignaut, D. (2019). Alkaline treatment for preventing acidosis in the rumen culture fermenting carbohydrates: An experimental study in vitro. Journal of Advanced Veterinary and Animal Research, 6(1): 100-107. https://doi.org/10.5455/javar.2019.f319
[20] Grant, R.J., Ferraretto, L.F. (2018). Silage review: Silage feeding management: Silage characteristics and dairy cow feeding behavior. Journal of Dairy Science, 101(5): 4111-4121. https://doi.org/10.3168/jds.2017-13729
[21] Ntakyo, P.R., Kirunda, H., Tugume, G., Natuha, S. (2020). Dry season feeding technologies: assessing the nutritional and economic benefits of feeding hay and silage to dairy cattle in South-Western Uganda. Open Journal of Animal Sciences, 10(3): 627-648. https://doi.org/10.4236/ojas.2020.103041
[22] Wilkinson, J.M., Rinne, M. (2018). Highlights of progress in silage conservation and future perspectives. Grass and Forage Science, 73(1): 40-52. https://doi.org/10.1111/gfs.12327
[23] Adesogan, A.T., Newman, Y.C. (2010). Silage harvesting, storing, and feeding. University of Florida IFAS Extension SSAGR-177.
[24] Muck, R.E., Kung Jr, L., Collins, M. (2020). Silage production. Forages: The Science of Grassland Agriculture, 2: 767-787.
[25] Kung Jr, L. (2010). Understanding the biology of silage preservation to maximize quality and protect the environment. In Proceedings, 2010 California Alfalfa & Forage Symposium and Corn/Cereal Silage Conference, pp. 1-2.
[26] Okoye, C.O., Wang, Y., Gao, L., Wu, Y., Li, X., Sun, J., Jiang, J. (2023). The performance of lactic acid bacteria in silage production: A review of modern biotechnology for silage improvement. Microbiological Research, 266: 127212. https://doi.org/10.1016/j.micres.2022.127212
[27] Alba, H.D.R., Lima, V.G.O., Carvalho, S.T., Ítavo, L.C.V., Pinto, L.F.B., Carneiro, P.L.S., Oliveira, R.L. (2023). Rumen content from slaughterhouse as an alternative inoculum source for in vitro analysis of feeds: A multivariate approach. Ruminants, 3(4): 360-372. https://doi.org/10.3390/ruminants3040030
[28] Darwin, D. (2016). Anaerobic co-digestion of cocoa husk with digested swine manure: Evaluation of biodegradation efficiency in methane productivity. Agricultural Engineering International: CIGR Journal, 18(4): 147-156.
[29] Darwin, Cheng, J.J., Gontupil, J., Liu, Z. (2016). Influence of total solid concentration for methane production of cocoa husk co-digested with digested swine manure. International Journal of Environment and Waste Management, 17(1): 71-90. https://doi.org/10.1504/IJEWM.2016.076432
[30] Bolsen, K.K., Ashbell, G., Weinberg, Z.G. (1996). Silage fermentation and silage additives-review. Asian-Australasian Journal of Animal Sciences, 9(5): 483-493. https://doi.org/10.5713/ajas.1996.483
[31] Rose-Dye, T.K., Burciaga-Robles, L.O., Krehbiel, C.R., Step, D.L., Fulton, R.W., Confer, A.W., Richards, C.J. (2011). Rumen temperature change monitored with remote rumen temperature boluses after challenges with bovine viral diarrhea virus and Mannheimia haemolytica. Journal of Animal Science, 89(4): 1193-1200. https://doi.org/10.2527/jas.2010-3051
[32] Darwin, Thifal, M., Alwi, M., Murizal, Z., Pratama, A., Rizal, M. (2023). The synthesis of biodiesel from palm oil and waste cooking oil via electrolysis by various electrodes. Case Studies in Chemical and Environmental Engineering, 8: 100512. https://doi.org/10.1016/j.cscee.2023.100512
[33] Darwin, Pratama, A., Mardhotillah. (2021). Biodiesel production from waste frying oils by potassium methoxide supported on activated carbon catalysts from lignocellulosic biomass. Acta Technologica Agriculturae, 24(4): 153-158. https://doi.org/10.2478/ata-2021-0026
[34] Björnsson, L., Murto, M., Jantsch, T.G., Mattiasson, B. (2001). Evaluation of new methods for the monitoring of alkalinity, dissolved hydrogen and the microbial community in anaerobic digestion. Water Research, 35(12): 2833-2840. https://doi.org/10.1016/S0043-1354(00)00585-6
[35] Darwin, Muliawati, A., Bulan, R. (2022). Performance of mixed-microbial culture from civet fecal suspensions on physicochemical composition of wet fermented Arabica coffee. Current Research in Nutrition and Food Science, 10(3): 918-927. https://doi.org/10.12944/CRNFSJ.10.3.9
[36] Kodama, T., Ichikawa, T., Hidaka, K., Furuya, K. (2015). A highly sensitive and large concentration range colorimetric continuous flow analysis for ammonium concentration. Journal of Oceanography, 71: 65-75. https://doi.org/10.1007/s10872-014-0260-6
[37] Wu, D., Xia, Q., Zhou, J., Ye, X. (2022). Evaluation of water ammonium ion test kit and its feasibility for the analysis of protein in food. Molecules, 27(15): 4689. https://doi.org/10.3390/molecules27154689
[38] Thiex, N.J., Manson, H., Anderson, S., Persson, J.Å., Collaborators. (2002). Determination of crude protein in animal feed, forage, grain, and oilseeds by using block digestion with a copper catalyst and steam distillation into boric acid: collaborative study. Journal of AOAC International, 85(2): 309-317. https://doi.org/10.1093/jaoac/85.2.309
[39] AOAC International. (2009). AOAC Official Method 2001.11 Protein (crude) in animal feed, forage (plant tissue), grain and oilseeds. Official Methods of Analysis of AOAC International.
[40] Horwitz, W. (2000). AOAC official method 962.09. Fiber (crude) in animal feed and pet food. Official Methods of Analysis of AOAC International.
[41] Darwin, Pratama, A., Mardhotillah. (2021). Organic waste conversion via continuous anaerobic co-digestion of oil palm empty fruit bunches and cow manure: Evaluation of feeding regime on methane production. Acta Technologica Agriculturae, 24(1): 8-13. https://doi.org/10.2478/ata-2021-0002
[42] Darwin, Triovanta, U., Rinaldi, R. (2019). Two-stage anaerobic co-digestion of landfill leachate and starch wastes using anaerobic biofilm reactor for methane production. Progress in Agricultural Engineering Sciences, 15(1): 53-70. https://doi.org/10.1556/446.15.2019.1.4
[43] Rice, E.W., Baird, R.B., Eaton, A.D. (2017). Standard Methods for the Examination of Water and Wastewater 23rd Edition. American Public Health Association (APHA), Washington, D.C., U.S.A.
[44] Darwin, Triovanta, U., Rinaldi, R., Pratama, A. (2019). Anaerobic acidification of coconut water waste by Lactobacillus acidophilus culture for biotechnological production of lactic acid. Acta Universitatis Agriculturae et Silviculturae Mendelianae Brunensis, 67(6): 1433-1440. https://doi.org/10.11118/actaun201967061433
[45] Lützhøft, H.C.H., Boe, K., Fang, C., Angelidaki, I. (2014). Comparison of VFA titration procedures used for monitoring the biogas process. Water Research, 54: 262-272. https://doi.org/10.1016/j.watres.2014.02.001
[46] Sun, H., Wu, S., Dong, R. (2016). Monitoring volatile fatty acids and carbonate alkalinity in anaerobic digestion: Titration methodologies. Chemical Engineering & Technology, 39(4): 599-610. https://doi.org/10.1002/ceat.201500293
[47] Darwin, D. (2019). Rapid determination of lactic acid in anaerobic biological treatment process using a portable sensitive lactate biosensor. BioTechnologia. Journal of Biotechnology Computational Biology and Bionanotechnology, 100(2): 115-120. https://doi.org/10.5114/bta.2019.85320
[48] Darwin. (2019). Determination of glucose concentration in anaerobic acidification cultures by portable glucose monitoring system. Asian Journal of Chemistry, 31(4): 763-766.
[49] Rustomo, B., AlZahal, O., Cant, J.P., Fan, M.Z., Duffield, T.F., Odongo, N.E., McBride, B.W. (2006). Acidogenic value of feeds II. Effects of rumen acid load from feeds on dry matter intake, ruminal pH, fibre degradability and milk production in the lactating dairy cow. Canadian Journal of Animal Science, 86(1): 119-127. https://doi.org/10.4141/A04-075
[50] Ryu, C.H., Bang, H.T., Lee, S., Kim, B., Baek, Y.C. (2022). Effects of feed composition in different growth stages on rumen fermentation and microbial diversity of Hanwoo steers. Animals, 12(19): 2606. https://doi.org/10.3390/ani12192606
[51] Satter, L.D., Slyter, L.L. (1974). Effect of ammonia concentration on rumen microbial protein production in vitro. British Journal of Nutrition, 32(2): 199-208. https://doi.org/10.1079/BJN19740073
[52] Dewhurst, R.J., Newbold, J.R. (2022). Effect of ammonia concentration on rumen microbial protein production in vitro. British Journal of Nutrition, 127(6): 847-849. https://doi.org/10.1079/BJN19740073
[53] Selim, A.S.M., Hasan, M.N., Rahman, M.A., Rahman, M.M., Islam, M.R., Bostami, A.R., Islam, S., Tedeschi, L.O. (2022). Nutrient content and in vitro degradation study of some unconventional feed resources of Bangladesh. Heliyon, 8(5): e09496. https://doi.org/10.1016/j.heliyon.2022.e09496
[54] Tachie, C.Y., Onuh, J.O., Aryee, A.N. (2024). Nutritional and potential health benefits of fermented food proteins. Journal of the Science of Food and Agriculture, 104(3): 1223-1233. https://doi.org/10.1002/jsfa.13001
[55] McDonald, P., Edwards, R.A., Greenhalgh, J.F.D., Morgan, C.A., Sinclair, L.A., Wilkinson, R.G. (2010). Animal Nutrition Seventh Edition. Pearson. UK.
[56] Valente, T.N.P., Sampaio, C.B., Lima, E.D.S., Deminicis, B.B., Cezário, A.S., Santos, W.B.R.D. (2017). Aspects of acidosis in ruminants with a focus on nutrition: A review. Journal of Agricultural Science, 9(3): 90. https://doi.org/10.5539/JAS.V9N3P90
[57] Darwin, Humaira, T., Muliawati, A. (2022). Identification and characterization of acidosis on in vitro rumen fermentation with feeds based on grass, rice bran, concentrate, and tofu pulp. Journal of Applied Biology & Biotechnology, 10: 53-58. https://doi.org/10.7324/JABB.2022.10s108
[58] Darwin, Mahyudi, R., Sundari, D., Muliawati, A. (2021). Influence of solid-state fermentation on Robusta coffee (Coffea cannephora) beans inoculated with microbial culture of civet fecal suspensions. Research on Crops, 22(4): 968-976. https://doi.org/10.31830/2348-7542.2021.158
[59] Moosavi, M. (2017). Bentonite clay as a natural remedy: A brief review. Iranian Journal of Public Health, 46(9): 1176.
[60] EFSA Panel on Additives and Products or Substances used in Animal Feed (FEEDAP). (2011). Scientific Opinion on the safety and efficacy of bentonite (dioctahedral montmorillonite) as feed additive for all species. EFSA Journal, 9(2): 2007. https://doi.org/10.2903/j.efsa.2011.2007
[61] Seheult, J., Fitzpatrick, G., Boran, G. (2017). Lactic acidosis: An update. Clinical Chemistry and Laboratory Medicine (CCLM), 55(3): 322-333. https://doi.org/10.1515/cclm-2016-0438
[62] Stojanović, B., Grubić, G., Adamović, M., Radivojević, M., Šamanc, H. (2009). Effect of bentonite in pelleted feed for calves. In 1st Workshop Feed-to-Food FP7 REGPOT-3. XIII Symposium Feed Technology, Proceedings, Novi Sad, Serbia, pp. 62-167.
[63] Addy, K., Green, L., Herron, E. (2004). pH and Alkalinity. University of Rhode Island, Kingston.
[64] De Lima, C.J., Coelho, L.F., Contiero, J. (2010). The use of response surface methodology in optimization of lactic acid production: focus on medium supplementation, temperature and pH control. Food Technology and Biotechnology, 48(2): 175-181. https://hrcak.srce.hr/53626
[65] Khalid, K. (2011). An overview of lactic acid bacteria. International Journal of Biosciences, 1(3): 1-13.
[66] Li, S., Danscher, A.M., Plaizier, J.C. (2013). Subactue Ruminal Acidosis (SARA) in dairy cattle: New developments in diagnostic aspects and feeding management. Canadian Journal of Animal Science, 94(1): 353-364.
[67] Abdela, N. (2016). Sub-acute ruminal acidosis (SARA) and its consequence in dairy cattle: A review of past and recent research at global prospective. Achievements in the Life Sciences, 10(2): 187-196. https://doi.org/10.1016/j.als.2016.11.006
[68] Damato, A. (2023). Safe use of bentonite in dairy cows. assessment of physiological responses by studying metabolome and mineral content in biofluids and milk characteristics. Universita Degli Studi Di Padova. Italy.
[69] Eturki, S., Ayari, F., Jedidi, N., Ben Dhia, H. (2012). Use of clay mineral to reduce ammonium from wastewater. Effect of various parameters. Surface Engineering and Applied Electrochemistry, 48(3): 276-283. https://doi.org/10.3103/S1068375512030064
[70] He, L., Wu, H., Wang, G., Meng, Q., Zhou, Z. (2018). The effects of including corn silage, corn stalk silage, and corn grain in finishing ration of beef steers on meat quality and oxidative stability. Meat Science, 139: 142-148. https://doi.org/10.1016/j.meatsci.2018.01.023
[71] Erdman, R.A. (1988). Dietary buffering requirements of the lactating dairy cow: A review. Journal of Dairy Science, 71(12): 3246-3266. https://doi.org/10.3168/jds.S0022-0302(88)79930-0