Samanhudi*![]() | Andriyana Setyawati
| Andriyana Setyawati![]() | Muji Rahayu
| Muji Rahayu![]() | Iswahyudi
| Iswahyudi![]() | Sabitha R. Dewanti
| Sabitha R. Dewanti![]() | Alfida Muthi’ah
| Alfida Muthi’ah![]()
© 2024 The authors. This article is published by IIETA and is licensed under the CC BY 4.0 license (http://creativecommons.org/licenses/by/4.0/).
OPEN ACCESS
Palmarosa plant (Cymbopogon martinii) comes from East India and Turkey, in Indonesia palmarosa is often called palmarosa grass which belongs to the Poaceae family. Palmarosa produces essential oil known as palmarosa oil. This research was carried out in a greenhouse which is located at Rumah Atsiri Indonesia, Tawangmangu, Karanganyar, from July to November 2023. This research aims to obtain the interaction of water availability and arbuscular mycorrhiza towards growth and result of palmarosa essential oil, to obtain the best level of water availability on growth and the results of palmarosa essential oil, to get the best dose of arbuscular mycorrhiza on the growth and result of palmarosa essential oil. The research was carried out using a factorial pattern with Completely Randomized Design (CRD) consisting of 2 factors. The first factor is the administration of arbuscular mycorrhiza which consists of four levels, namely, without mycorrhiza (control), mycorrhiza 10 g polybag-1, mycorrhiza 20 g polybag-1 and mycorrhiza 30 g polybag-1. The second factor is water availability which consists of field capacity of 100%, 75%, 50% and 25%. Each treatment was repeated 5 times. Data were analyzed using analysis of variance if the F-count was greater than the F-table, followed by the DMRT test at the 5% level. The results showed that giving both treatments did not show significant results. The application of arbuscular mycorrhiza was very significant in the parameters of plant height and stem diameter. The field capacity treatment at various levels was very significant for the parameters of plant height, stem diameter, number of tillers and number of leaves.
aromatic crops, essential oil, drought stress, geraniol, organic fertilizer
Essential oils are among the products of plantation commodities with high export value. They possess a high selling value due to their ability to produce fragrances containing volatile compounds. These volatile compounds in essential oils have been studied to provide various benefits such as antimicrobial, anti-inflammatory, and aromatherapy properties [1]. Essential oils mostly consist of terpenoids with isomers that include positional, geometric, spatial, and optical variations [2]. Essential oils are found in many aromatic plants and contribute significantly to the food, pharmaceutical, and cosmetic industries. The palmarosa plant is one of the sources of essential oils that has high potential in Indonesia, producing an essential oil known as palmarosa oil.
The palmarosa plant (Cymbopogon martinii) originates from East India and Turkey; in Indonesia, it is often referred to as palmarosa grass and belongs to the Poaceae family. The essential oil produced by palmarosa is commonly used in aromatherapy due to its fragrance and also serves as an antioxidant and antimicrobial agent. Palmarosa is highly valued for its essential oils found in its leaves and inflorescences, with geraniol being the main constituent (75-90%) of the oil [3].
The increase content in geraniol in palmarosa is a response to the formation of secondary metabolites. The level of plant tolerance to various abiotic environmental conditions influences the secondary metabolite content of palmarosa. Several factors in the maintenance of palmarosa plants can increase both production and the quality of the contained oil. According to research [4], the addition of manure fertilizer and lemongrass compost can increase the weight per clump by 179.19% and the geraniol content by 81.04%. Manure contains many organic materials that can making them more fertile and increasing the diversity of soil microorganisms [5]. The fertilizers as providers of macro and micronutrients [6, 7], enhance the volume of plant roots [8], and increase the value of pH and organic materials of soil [9-11].
The factors that influence the absorption of organic matter in the soil are moisture, temperature and soil aeration [12]. The growth response of palmarosa plants is also related to the nutrient absorption mechanism by plants, both macro and micronutrients which are generally given in the form of compounds [13]. Organic compounds and mineral nutrients are also important for plant growth and development [14] with one of the roles being played by Potassium ions (K+). The provision of K+ ions can stimulate the photosynthesis process and produce more photosynthate, resulting in increased palmarosa production and yield. Gusmaini and Syakir [15] conducted research on the Cymbopogon family, specifically Cymbopogon nardus, and found that the application of Potassium at a rate of 30 kg/Ha could yield oil at 1.20-1.28%, oil content at 1.40-1.70%, and increase potassium nutrient uptake by 11.76%.
In addition to those factors, water availability for plants also plays a significant role in the growth and essential oil content of palmarosa plants. When plants experience water stress, either due to drought or excess water, they can produce higher levels of secondary metabolites as a defensive response. Water availability is the primary factor for plant growth, and it must align with the plant's needs. The soil's ability to store water optimally is termed field capacity, which is the capacity to hold water after applying water until it reaches the saturation point. The value of field capacity varies significantly depending on the soil type [16].
It is necessary to control this stress condition to prevent the plant from dying, one way being the addition of mycorrhizal biological agents. Plants supplemented with mycorrhizae have high tolerance to both biotic and abiotic stress conditions, including water stress [17]. Maintaining water at a specific level and introducing arbuscular mycorrhiza can increase the number of plant leaves [18]. The limited research discussing efforts to enhance the quality of palmarosa essential oil in Indonesia warrants attention within the essential oil industry sector. Cultivation methods involving water availability and the application of arbuscular mycorrhiza in andosol soil have not been put into practice. Hence, it is necessary to conduct research to determine palmarosa's response to varying water availability and the administration of arbuscular mycorrhiza.
This research was conducted from July to November 2023 at Rumah Atsiri Indonesia, located on Watusambang Street in Watusambang, Plumbon, Tawangmangu District, Karanganyar Regency, Central Java, Indonesia. The research followed a factorial pattern in a Completely Randomized Design (CRD) consisting of 2 factors. The first factor was the administration of arbuscular mycorrhiza, which had four levels: without mycorrhiza (control), mycorrhiza 10 g polybag-1, mycorrhiza 20 g polybag-1, and mycorrhiza 30 g polybag-1. The types of arbuscular mycorrhizal fungi included are Glomus claroideum, Glomus fasisculatum, Glomus moseae, Glomus etunicatum, Acaulospora rogusa, and Acaulospora tuberculata. The second factor was water availability, with field capacity levels of 100%, 75%, 50%, and 25%. The calculation of water volume for irrigation is based on the field capacity of the soil used.
The soil in polybags measuring 45 × 45 cm has a weight of 15 kg and a field capacity of 3 liters. Each treatment was replicated 5 times. The planting material used was palmarosa seeds (C. martini) obtained from Rumah Atsiri Indonesia. Palmarosa seedlings are grown for 2 months until the plants reach the same height, approximately 15-20 cm. The planting medium used is andosol soil obtained from Rumah Atsiri Indonesia, which has been loosened. Arbuscular mycorrhiza was applied during transplanting, one week before the water availability treatment. Palmarosa is planted in polybags measuring 45 × 45 cm with a planting distance between polybags of 50 × 50 cm. The maintenance given to these plants includes watering every 6 days with a volume corresponding to the treatment level, weed management, and pest control if present on the plants.
Data collection involved observation, measurement, and documentation. The observation parameters included plant height, stem diameter, number of tillers, and number of leaves, measured using a tape measure, digital caliper, and counting in one clump, respectively. The data analysis method used was Analysis of Variance (ANOVA) with the F-test at a 5% significance level. If a significant difference was found, Duncan's Multiple Range Test (DMRT) at the 5% level was conducted. To carry out this study, a Completely Randomized Design (CRD) was employed and organized in a factorial design comprising 2 treatment factors. Each treatment was replicated five times, resulting in 16 treatment combinations per replication. Normality of residuals and homogeneity were verified with Kolmogorov-Smirnov’s and Levene’s tests, and normality was achieved, when necessary, by transforming growth variables (Log10). All data has been analyzed and obtained normal and homogeneous results.
Before carrying out further tests, the data was analyzed for normality and homogeneity which are presented as follows (Table 1).
Table 1. Result of normality and homogeneity test to all growth variables at 28 days after planting (DAP)
|
Variables |
Normality Testa |
Homogeneity Testb |
|
Plant heightc |
Sig. 0.940 |
Sig. 0.088 |
|
Stem Diameter |
Sig. 0.196 |
Sig. 0.122 |
|
No. of tillersc |
Sig. 0.056 |
Sig. 0.069 |
|
No. of leavesc |
Sig. 0.379 |
Sig. 0.075 |
Note: aKolmogorov-Smirnov’s test significance level is > 0.05; bLevene’s test significance level is > 0.05; cData were analyzed under log transformation to retain the hypothesis.
Based on Table 1, the results of the normality test and homogeneity test using the SPSS application obtained significant Kolmogrov-Smirnov's with significance values > 0.05 for plant height, stem diameter, number of tillers and number of leaves. Thus, the results of the data in the table are normally distributed. In the homogeneity test, it was found that the significance values > 0.05 for all observed variables, this shows that the data is homogeneous and further test can be carried out.
The addition of mycorrhiza and watering at different levels can result in different growths for palmarosa. Excessive water levels can cause plants to rot, while insufficient water can lead to wilting and hindered growth. The presence of mycorrhiza in symbiosis with plant roots plays a crucial role in plant growth, including enhancing nutrient uptake.
Based on Table 2, the observation parameters of palmarosa growth at 28 days after planting (DAP), including plant height, stem diameter, number of tillers, and number of leaves, in the treatments involving mycorrhiza and different levels of water availability, showed significant results. Each treatment had a significant effect on the parameters of plant height and stem diameter at the 1% level. Mycorrhiza treatment did not significantly affect the parameters of the number of tillers and number of leaves, but water availability had a significant effect on all parameters at the 1% level. This is different from the findings of the study [19], which applied arbuscular mycorrhiza to Foeniculum vulgare L. plants and were able to produce the highest number of leaves compared to other treatments, totalling 188.19 leaves. Based on the analysis results, it is evident that the interaction between mycorrhizal treatments and water availability does not significantly affect all parameters.
Table 2. Results of analysis of variance of palmarosa growth variables at 28 days after planting (DAP)
|
Treatment |
Plant Height |
Stem Diameter |
Number of Tillers |
Number of Leaves |
|
Mycorrhiza |
** |
** |
ns |
ns |
|
Water Availability |
** |
** |
** |
** |
|
Interaction |
ns |
ns |
ns |
ns |
Note: **: significant at 1% level. ns: non-significant.
Based on Table 3, the data were analyzed using the Duncan Multiple Range Test (DMRT) at the 5% level. It shows that mycorrhizal treatment has a significant effect on plant height and stem diameter (Figure 1). Based on the treatment of 10 g polybag-1 mycorrhiza, it increased plant height to 68.49 cm, but this value was not significantly different from the control treatment or without giving mycorrhiza. Mycorrhiza treatment increased plant height and stem diameter, although not significantly. However, excessive mycorrhizal application is also not economically efficient and can hinder plant growth. The research results indicate that applying mycorrhiza at 10 g polybag-1 resulted in taller plants and larger stem diameters compared to applying 20 g and 30 g polybag-1. This is consistent with the study [20], which found that excessively high doses of mycorrhiza inhibit plant growth, including plant height, stem diameter, and wet weight. Mycorrhiza aids in the absorption of P ions beyond the reach of plant roots and converts P into primary or secondary P ions with the help of the phosphatase enzyme. According to study [21], mycorrhiza enhances P nutrient uptake in plant height, stem diameter, and flower formation time, and increases P nutrient uptake in upland rice plant leaf tissue by an average of 0.26%. The treatment of mycorrhiza at 20 g polybag-1 was not significantly different from mycorrhiza at 30 g polybag-1; both levels resulted in a lower average plant height compared to not using mycorrhiza and using mycorrhiza at 10 g polybag-1 (Figure 2).
Table 3. Effect of mycorrhizal levels on plant height and stem diameter of palmarosa (C. martinii) at 28 days after planting (DAP)
|
Treatment (g polybag-1) |
Plant Height (cm) |
Stem Diameter (mm) |
|
0 Mycorrhiza |
67.48ab |
4.12a |
|
10 Mycorrhiza |
68.49a |
3.94ab |
|
20 Mycorrhiza |
51.42c |
3.56bc |
|
30 Mycorrhiza |
57.10bc |
3.29c |
Note: Numbers in columns followed by the same letter indicate that they are not significantly different at the Duncan's Multiple Range Test at the 5% level.
Figure 1. Graph of average palmarosa plant height at 28 DAP
Figure 2. Palmarosa plants with treatment of (a) control (b) 20 g polybag-1 mycorrhiza
The mycorrhizal treatment affected the stem diameter of palmarosa plants. The treatment without mycorrhiza was not significantly different from the treatment with mycorrhiza in 10 g polybag-1, with average stem diameters of 4.12 mm and 3.94 mm, respectively. However, both levels produce a relatively large average stem diameter. The dose mycorrhiza of 10 g polybag-1 was significantly different from 30 g polybag-1. The additional dose of mycorrhiza 30 g polybag-1 could reduce the stem diameter by 3.29 mm. This is because the dose of mycorrhiza is too high. During the dry season, high spore densities and lower root colonization will occur [22]. High colonization of many spores provided will indicate competition for the space where symbiosis occurs [23] so that there may be a decline at the end of the observation. Based on Figure 3, it can be seen that increasing the level of mycorrhiza application does not necessarily increase plant growth in terms of stem diameter. However, providing mycorrhiza at the 10 g polybag-1 and 20 g polybag-1 levels has been proven to increase plant growth under low water availability (25% field capacity). Arbuscular mycorrhiza can help maintain plant growth under less-than-optimal environmental conditions by expanding the area of water and nutrient absorption. According to study [24], mycorrhiza is useful for improving soil structure by increasing organic matter and the soil's ability to maintain groundwater. Begum et al. [25] confirmed that arbuscular mycorrhizal fungi have extensive hyphae that can enhance water and mineral absorption in plants under abiotic stress conditions. This aligns with the results of a study by Putra et al. [26] on dwarf napier grass plants (Pennisetum purpureum cv. Mott) or purple elephant grass, where the application of arbuscular mycorrhizal fungi improved and enhanced the growth of dwarf napier grass plants in abiotic stress environments. The role of arbuscular mycorrhiza is to optimize nutrient and water absorption, leading to greater plant height and stem diameter compared to plants without mycorrhiza. The benefits of providing mycorrhiza are the same as adding organic fertilizer to plants, according to research by Rahayu et al. [27], similar to providing organic fertilizer, in soybean plants it increases the growth of local soybean plants, including plant height, number of leaves, and leaf area.
Figure 3. Graph of average stem diameter (mm) at 28 DAP
According to the results of the DMRT test, it is clear that the treatment involving water availability has a significant impact on the observed variables. These variables include plant height, stem diameter, number of tillers, and number of leaves. This indicates that varying levels of water availability can greatly influence the growth characteristics. The water availability treatment at 100% field capacity (FC) level is significantly different from the 50% treatment, where at 100% water availability the average plant height is relatively high, namely 83.46 cm and decreases at 25% water availability with a value of 55.15 cm (Figure 4). Water is an important factor in plant growth. According to Riyadi and Siswanti [28], water has an important role in cell enlargement and elongation, regulating cell turgor pressure, and is an active component of meristem tissue in carrying out physiological processes in plants. Table 4 also indicates that the water availability treatment at the 50% FC level is significantly different from the treatment at 25% FC; both levels produce the lowest average plant height compared to the other treatment levels. Severe drought stress (25% field capacity) has a considerable impact on plant height because, as noted by Barickman et al. [29], stress treatments can hinder plant growth and development, disrupt metabolic processes, and lead to impaired cell division and elongation compared to plants with adequate water treatment. Insufficient water availability can also result in moisture stress. According to Fadiji et al. [30], low watering reduces cell water potential, leading to stomatal closure that affects cell growth and elongation. Numerous plants, such as corn, rice, wheat, and barley, have been extensively studied, revealing that drought stress conditions can impede plant growth and disrupt flowering during the reproductive stage, ultimately causing yield losses.
Figure 4. Palmarosa plants with water availability treatment of (a) 100% field capacity (b) 25% field capacity
Table 4. Effect of water availability level on plant height, stem diameter, number of tillers, and number of leaves of palmarosa (C. martinii) at 28 DAP
|
Treatment (%) |
Plant Height (cm) |
Stem Diameter (mm) |
Number of Tillers |
Number of Leaves |
|
100 FC |
83.46a |
4.53a |
60.45a |
221.40a |
|
75 FC |
63.74b |
3.81b |
58.60ab |
203.20ab |
|
50 FC |
55.15b |
3.56b |
51.05bc |
180.00bc |
|
25 FC |
42.14c |
3.01c |
43.80c |
151.60c |
Note: Numbers in columns followed by the same letter indicate that they are not significantly different at the Duncan's Multiple Range Test at the 5% level.
Based on Table 4, the stem diameter shows that the 100% water availability treatment gives the largest average stem diameter and is significantly different from the water availability levels of 50 and 25%. This is consistent with the research by Nawaz et al. [31], which found that providing no stress water to plants resulted in maize plants with more optimal growth compared to stress water treatments. Plants with no stress water had taller plants, larger stem diameters, greater leaf area, longer roots, and higher plant fresh biomass. Each level gives an average stem diameter of 4.53; 3.81; 3.56 and 3.01 mm. 100% water availability increased palmarosa stem diameter by 4.53 mm and decreased when 25% water was given to 3.01 mm. These results showed that the need for water is an important limiting factor in plant growth. According to Arsa et al. [32], water is very helpful in increasing the volume of the stem because water circulates the results of photosynthesis and acts as a solvent for cell nuclei in the process of growth and development of the stem. Analysis conducted by Balducci et al. [33] also shows that low soil moisture can significantly reduce plant stem girth. Drought can limit plant growth by inhibiting meristematic tissue cell division, cell expansion in elongation areas, and driving complex changes in palisade tissue and sponge tissue during adaptation. Drought causes an imbalance in plant water status in the short term which increases the duration of stem contraction and reduces the duration of stem expansion and reduces growth during the growing season.
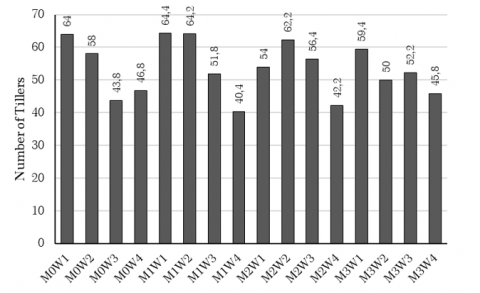
Table 4 also shows that the 100% FC water availability treatment is significantly different from the 25% FC water availability treatment in terms of the number of tillers variable. The highest average number of tillers was observed with the 100% FC water availability treatment, reaching 60.45 saplings per clump. Based on Figure 5, the number of tillers shows that the 10 grams of arbuscular mycorrhiza and 100% field capacity treatment gives the largest average, amounting to 64.4 tillers per clump at 28 DAP. The number of tillers per plant is the most important characteristic, followed by plant height and number of leaves. The greater the number of tillers, the more leaves the plant will have, leading to better yields for essential oil-producing plants. Increasing number of leaves also increase leaf area and produce more photosynthesis, which is crucial for the nutrient uptake, thus improving the overall quality and quantity of the yield. According to Nandapure et al. [34], the increase in the number of tillers is due to the availability of better and more balanced nutrients resulting in higher uptake of nutrients by plants and ultimately producing a greater number of tillers. However, the 50% FC treatment was not significantly different from the 25% FC treatment, both yielding a low average number of tillers, specifically 51.04 and 43.80 tillers per clump, respectively. According to Dwicaksono [35], reducing the number of tillers is an attempt by plants on dry land to use water efficiently. Optimizing the distribution of assimilation into a small number of tillers aims to reduce the number of tillers to reduce evaporation. Based on low levels of water, such as 50% and 25% FC, led to drought stress conditions in plants. According to Ahmad et al. [36], drought stress significantly impacts yield and yield-related traits, including tiller number, grain production, and straw production. Drought stress can reduce water potential in the soil, affecting plant cells' ability to absorb nutrients. Generally, plants decrease phosphorus (P) uptake under water deficit conditions. Drought can hinder P absorption by limiting P distribution to the roots and affecting other water-related factors in plants under drought stress, resulting in suboptimal growth compared to plants receiving adequate water. Additionally, according to Herawati [18], the water treatment level did not significantly affect the number of tillers.
Figure 5. Graph of the average number of palmarosa tillers at 28 DAP
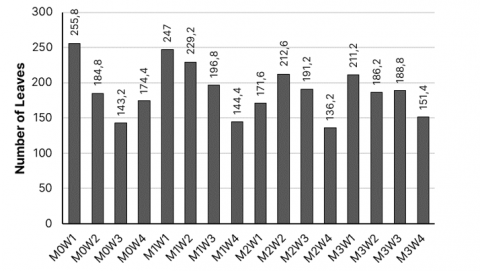
Table 4 shows that the water availability treatment at the 100% FC level is significantly different from the treatments at and 50% FC in terms of the number of leaves variable. The highest average number of leaves was observed at the 100% FC level, specifically 221.40 leaves per cluster. Based on Figure 6, the number of leaves shows that the 0 grams of arbuscular mycorrhiza and 100% field capacity treatment gives the largest average, amounting to 255.8 leaves per cluster at 28 DAP. This had an effect because the plant's water needs were met optimally, thereby increasing the number of leaves, according to Azzahro et al. [37], when the need for photosynthesis is fulfilled, the amount of photosynthesis produced also increase. Part of the photosynthate produced will be distributed to determine the number of leaves. The higher the number of leaves, the higher the photosynthesis process, the higher the photosynthate produced. The number of leaves indicates the nitrogen content in the plant, N supply increases chlorophyll content, total protein, sugar content, protein, fat, auxin formation stimulant to soften the cell walls, improve plant's ability in absorbing water, and metabolites associated with photosynthesis [38-41]. On the other hand, the 50% and 25% FC treatments did not show significant differences between them. The 25% FC level resulted in a relatively low average number of leaves compared to other levels of 151.60 leaves per hill. According to Evlin et al. [42], for plants, water is needed for the processes of photosynthesis and transpiration (evaporation of water from the surface of plant leaves), transporting and dissolving nutrients from the soil, water being the main element of plants. Therefore, if water intake is insufficient, plant growth will be adversely affected because the rate of water absorption cannot keep up with the speed of water evaporation from the plant. This imbalance results in disrupted physiological processes, leading to stunted and abnormal growth. Plants deprived of adequate water from the beginning of their growth phase will experience inhibited development and exhibit abnormal growth patterns. The rate of transpiration tends to increase as the volume of water provided to the plant is elevated. This relationship highlights that with higher watering volumes, plants experience a more significant loss of water through the process of transpiration, leading to a higher overall rate. According to Riyadi and Siswanti [28], an increase in transpiration rate leads to a higher rate of photosynthesis in plants because the opening and closing of stomata are controlled by turgor pressure from guard cells, and CO2 diffusion occurs more effectively compared to plants under drought stress. If the stomata are closed, CO2 fixation decreases, and the gas exchange process is hindered. Consequently, plants cannot carry out photosynthesis optimally to produce energy sources for generating new leaf shoots.
Figure 6. Graph of the number of palmarosa leaves at 28 DAP
Note: M0: 0 gram of arbuscular mycorrhiza. M1: 10 grams of arbuscular mycorrhiza. M2: 20 grams of arbuscular mycorrhiza. M3: 30 grams of arbuscular mycorrhiza. W1: 100% field capacity. W2: 75% field capacity. W3: 50% field capacity. W4: 25% field capacity.
The treatment involving water availability and the application of arbuscular mycorrhiza significantly affected the vegetative growth of palmarosa (C. martinii). Specifically, a water availability of 100% field capacity (FC) showed a notable impact on plant height, stem diameter, number of tillers, and number of leaves, leading to higher growth compared to other treatments. The use of arbuscular mycorrhiza, at a dose of 10 g per polybag, notably increased plant height. Furthermore, the 100% field capacity water treatment was the most effective, producing the highest values for plant height, number of tillers, number of leaves, and stem diameter. Interestingly, this treatment showed an equally response from the 75% field capacity treatment in terms of these growth parameters such as number of tillers and number of leaves, suggesting that using 75% field capacity can be a water-saving alternative without compromising plant growth significantly.
The authors sincerely thank the Directorate of Research, Technology, and Community Services (DRTPM) under the Ministry of Education, Culture, Research, and Technology Republic of Indonesia for offering financial support for this research through the Fundamental Research scheme in the fiscal year 2023.
[1] Mahfud, M., Ma'sum, Z., Bhuana, D.S., Altway, A., Yuniati, Y. (2022). A comparison of essential oil extraction from the leaves of lemongrass (Cymbopogon nardus L.) using two microwave-assisted methods. Journal of Applied Engineering Science, 20(3): 881-888. https://doi.org/10.5937/jaes0-34006
[2] Khan, S., Sahar, A., Tariq, T., Sameen, A., Tariq, F. (2023). Essential oils in plants: Plant physiology, the chemical composition of the oil, and natural variation of the oils (chemotaxonomy and environmental effects, etc.). In Essential Oils, pp. 1-36. https://doi.org/10.1016/B978-0-323-91740-7.00016-5
[3] Pankaj, U., Kurmi, A., Lothe, N.B., Verma, R.K. (2021). Influence of the seedlings emergence and initial growth of palmarosa (Cymbopogon martinii (Roxb.) Wats. var. Motia Burk) by arbuscular mycorrhizal fungi in soil salinity conditions. Journal of Applied Research on Medicinal and Aromatic Plants, 24: 100317. https://doi.org/10.1016/j.jarmap.2021.100317
[4] Suryani, E., Idris, H. (2021). Productivity of palmarosa plant (Cymbopogon martinii) in ultisol soil with giving some organic fertilizer. Journal Galung Tropika, 10(2): 234-239. https://doi.org/10.31850/jgt.v10i2.746
[5] Yunus, A., Pujiasmanto, B., Cahyani, V.R., Lestariana, D.S. (2017). The effect of arbuscular mycorrhiza and organic manure on soybean growth and nutrient content in Indonesia. Bulgarian Journal of Agricultural Science, 23(4): 596-603.
[6] Samanhudi, S., Yunus, A., Pujiasmanto, B. (2018). Budidaya organik kunyit pada kluster biofarmaka kabupaten karanganyar. Caraka Tani: Journal of Sustainable Agriculture, 33(1): 34-41. https://doi.org/10.20961/carakatani.v33i1.19112
[7] Samanhudi, Pujiasmanto, B., Sudadi, Putra, I.H., Mumtazah, H.M. (2018). The efficiency of mycorrhiza biofertilizer treatment to the growth and yield of soybean. IOP Conference Series: Earth and Environmental Science, 142(1): 012073. https://doi.org/10.1088/1755-1315/142/1/012073
[8] Prasetyo, L., Widyastuti, Y., Yunus, A., Samanhudi. (2018). The growth response of Artemisia annua L. to organic fertilizer type in lowland. IOP Conference Series: Earth and Environmental Science, 142(1): 012019. https://doi.org/10.1088/1755-1315/142/1/012019
[9] Muktiyanta, M.N.A., Samanhudi, Yunus, A., Pujiasmanto, B., Minardi, S. (2018). Effectiveness of cow manure and mycorrhiza on the growth of soybean. IOP Conference Series: Earth and Environmental Science, 142(1): 012065. https://doi.org/10.1088/1755-1315/142/1/012065
[10] Samanhudi, Pujiasmanto, B., Yunus, A., Supyani, Suntoro, Widijanto, H., Prabowo, S.M. (2017). The effect of manure and mycorrhiza application to the soil microbes biodiversity in terms of increasing soybean yield in marginal land in Indonesia. Bulgarian Journal of Agricultural Science, 23(6): 994-1003.
[11] Samanhudi, Sudadi, Yunus, A., Pujiasmanto, B., Mahasti, W. (2017). Pengaruh kascing dan mikoriza terhadap pertumbuhan dan hasil Tribulus terrestris. Agrotek Indonesia, 2(1): 1-6. https://doi.org/10.33661/jai.v2i1.711
[12] Dinda, W.P., Triharyanto, E., Samanhudi. (2020). Effects of mulch on growth and yield of garlic bulbils at various fertilizing doses. IOP Conference Series: Earth and Environmental Science, 423(1): 012033. https://doi.org/10.1088/1755-1315/423/1/012033
[13] Prabowo, H., Samanhudi, Yuniastuti, E., Yunus, A. (2018). Effects of media combination with concentration of ab-mix nutrient on growth of banana shoots on in vitro. Bulgarian Journal of Agricultural Science, 24(3): 404-410.
[14] Mardhikasari, S., Yunus, A., Samanhudi. (2020). Modification of media for banana in vitro propagation with foliar fertilizer and coconut water in cv. Rajabulu. Caraka Tani: Journal of Sustainable Agriculture, 35(1): 23-32. https://doi.org/10.20961/carakatani.v35i1.27756
[15] Gusmaini, G., Syakir, M. (2020). Effect of potassium on growth, yield and quality of citronella grass. Jurnal Penelitian Tanaman Industri, 26(1): 32-39.
[16] van Lier, Q.D.J. (2017). Field capacity, a valid upper limit of crop availability water. Agricultural Water Management, 193: 214-220. https://doi.org/10.1016/j.agwat.2017.08.017
[17] Rani, B., Jatttan, M., Kumari, N., Prashad, J., Kumari, A., Sharmam, K.D., Madan, S. (2024). Effect of drought stress on grain quality of wheat and its mitigation through arbuscular mycorrhiza fungi. Journal of Environmental Biology, 45(2): 218-225. http://doi.org/10.22438/jeb/45/2/MRN-1979
[18] Herawati, N. (2022). Response of growth citronella grass (Cymbopogon nardus L) by Acaulospora sp. mycorrizha and different water treatments in ultisol. JERAMI: Indonesian Journal of Crop Science, 5(1): 34-41. https://doi.org/10.25077/jijcs.5.1.34-41.2022
[19] Rasouli, F., Nasiri, Y., Hassanpouraghdam, M.B., Asadi, M., Qaderi, T., Trifa, A., Strzemski, M., Dresler, S., Szczepanek, M. (2023). Seaweed extract and arbuscular mycorrhiza co‑application afect the growth responses and essential oil composition of Foeniculum vulgare L. Scientific Reports, 13: 11902. https://doi.org/10.1038/s41598-023-39194-3
[20] Matondang, A.M., Syafruddin, Jumini. (2020). Effect of mycorrhizal biofertilizer type and dosage against growth and yield of chilli (Capsicum annum L.) on andosol soil valley in Aceh Besar. Jurnal Ilmiah Mahasiswa Pertanian, 5(2): 101-110. https://doi.org/10.17969/jimfp.v5i2.15025
[21] Sahur, A., Junaid, M. (2022). Impact of arbuscular mycorrhizae and gogo rice interaction against rice blas disease (Prycularia oryzae). Journal of Tropical Mycorrhiza, 1(2): 45-56. https://doi.org/10.58222/jtm.v1i2.29
[22] Kebede, T.G., Birhane, E., Meles, K., Gerima, Y.G.E.W. (2023). Arbuscular mycorrhizal fungal spore density and root colonization in cochineal-infested Opuntia ficus-indica plants in northern Ethiopia. Arthropod-Plant Interactions, 17: 839-854. https://doi.org/10.1007/s11829-023-10005-w
[23] García, Td.C.V., Zárate, G.H., Téllez, L.I.T., Bello, J.J.B. (2023). Endomycorrhizal fungi inoculation improves development of Stevia (Stevia rebaudiana Bertoni) plantlets during ex vitro acclimatization. Symbiosis, 90: 53-60. https://doi.org/10.1007/s13199-023-00919-7
[24] Nugroho, P.A., Widyastuti, Y., Samanhudi, Yunus. A. (2017). Growth response of Artemisia annua by effect of types and composition of organic fertilizer in lowland. Journal of Agricultural Science and Technology B, 7(5): 339-345. https://doi.org/10.17265/2161-6264/2017.05.006
[25] Begum, N., Qin, C., Ahanger, M.A., Raza, S., Khan, M.I., Ashraf, M., Ahmed, N., Zhang, L. (2019). Role of arbuscular mycorrhizal fungi in plant growth regulation: Implications in abiotic stress tolerance. Frontiers in Plant Science, 10: 1068. https://doi.org/10.3389/fpls.2019.01068
[26] Putra, B., Warly, L., Evitayani, E., Utama, B.P. (2022). The role of arbuscular mycorrhizal fungi in phytoremediation of heavy metals and their effect on the growth of Pennisetum purpureum cv. Mott on gold mine tailings in Muara Bungo, Jambi, Indonesia. Biodiversitas Journal of Biological Diversity, 23(1): 478-485. https://doi.org/10.13057/biodiv/d230151
[27] Rahayu, M., Purwanto, E., Setyawati, A., Sakya, A.T., Samanhudi, Yunus, A., Purnomo, D., Handoyo, G.C., Arniputri, R.B., Na'Imah, S. (2021). Growth and yield response of local soybean in the giving of various organic fertilizer. IOP Conference Series: Earth and Environmental Science, 905(1): 012028. https://doi.org/10.1088/1755-1315/905/1/012028
[28] Riyadi, A.D.R., Siswanti, D.U. (2022). Effect of alkaline and drought stress on growth and SOD content in basil plant (Ocimum americanum). Jurnal Biodjati, 7(1): 119-131. https://doi.org/10.15575/biodjati.v7i1.16680
[29] Barickman, T.C., Adhikari, B., Sehgal, A., Walne, C.H., Reddy, K.R. Gao, W. (2021). Drought and elevated carbon dioxide impact the morpho-physiological profile of Basil (Ocimum basilicum L.). Crops, 1(3): 118-128. https://doi.org/10.3390/crops1030012
[30] Fadiji, A.E., Santoyo, G., Yadav, A.N., Babalola, O.O. (2022). Efforts towards overcoming drought stress in crops: Revisiting the mechanisms employed by plant growth-promoting bacteria. Frontiers in Microbiology, 13: 962427. https://doi.org/10.3389/fmicb.2022.962427
[31] Nawaz, H., Akgün, I., Şenyiğit, U. (2024). Effect of deficit irrigation combined with Bacillus simplex on water use efficiency and growth parameters of maize during vegetative stage. BMC Plant Biology, 24: 135. https://doi.org/10.1186/s12870-024-04772-8
[32] Arsa, I.A., Dato, C.D., Kasim, M. (2023). The growth components and productivity results of three corn varieties (Zea mays L) against drought stress through the frequency of water application. Jurnal Agrisa, 12(2): 147-161.
[33] Balducci, L., Deslauriers, A., Rossi, S., Giovannelli, A. (2019). Stem cycle analyses help decipher the nonlinear response of trees to concurrent warming and drought. Annals of Forest Science, 76(3): 1-18. https://doi.org/10.1007/s13595-019-0870-7
[34] Nandapure, S.P., Wankhade, S.G., Imade, S.R., Mahajan, R.V., Doiphode, G.S. (2020). Growth parameters and herbage yield of java citronella as influenced by nutrient management under inceptisol. International Journal of Current Microbiology and Applied Sciences, 9(5): 1711-1720. https://doi.org/10.20546/ijcmas.2020.905.192
[35] Dwicaksono, G. (2023). Seedling height on the growth of vetiver (Vetiveria Zizanioides L.) with a test dose of NPK fertilizer 16: 16: 16. Jurnal Ilmiah Mahasiswa Pertanian, 3(1): 1-12. https://jurnalmahasiswa.umsu.ac.id/index.php/jimtani/article/view/2282/2361.
[36] Ahmad, A., Aslam, Z., Javed, T., Hussain, S., Raza, A., Shabbir, R., Tauseef, M. (2022). Screening of wheat (Triticum aestivum L.) genotypes for drought tolerance through agronomic and physiological response. Agronomy, 12(2): 287. https://doi.org/10.3390/agronomy12020287
[37] Azzahro, H.U., Juliandra, Y., Susi, N., Sari, V. I. (2023). Interaksi Kompos Sampah Pasar Dan Volume Pemberian Air Terhadap Pertumbuhan Tanaman Serai Wangi (Andropogon Nardus L). Jurnal Agrotela, 4(2): 85-91.
[38] Dawiyah, R.Y.A., Yunus, A., Samanhudi, Widiyastuti, Y., Widodo. (2018). Shading and vermicompost effect on growth and flavonoid content of tapak liman (Elephantopus scaber L.). IOP Conference Series: Earth and Environmental Science, 142(1): 012022. https://doi.org/10.1088/1755-1315/142/1/012022
[39] Permana, H.H., Widyastuti, Y., Samanhudi, Yunus, A. (2018). Response of Artemisia annua L. to shade and manure fertilizer application in lowland altitude. IOP Conference Series: Earth and Environmental Science, 142(1): 012043. https://doi.org/10.1088/1755-1315/142/1/012043
[40] Riyana, D., Widiyastuti, Widodo, H., Purwanto, E., Samanhudi. (2018). Effect of manure and plants spacing on yield and flavonoid content of Elephantopus scaber L. IOP Conference Series: Earth and Environmental Science, 142(1): 012038. https://doi.org/10.1088/1755-1315/142/1/012038
[41] Yunus, A., Samanhudi, Brahmanto, N., Widyastuti. Y. (2018). Artemisia annua respon to various types of organic fertilizer and dose in lowland. IOP Conference Series: Earth and Environmental Science, 142(1): 012021. https://doi.org/10.1088/1755-1315/142/1/012021
[42] Evlin, F., Nurcahyani, E., Sumardi, S., Wahyuningsih, S.W. (2020). Kajian tumbuhan rumput-rumputan yang tahan terhadap cekaman kekeringan dan tanah masam. Al-Hayat: Journal of Biology and Applied Biology, 10(1): 1-15.