Biofilm Formation and Gene Expression in Carbapenem-Resistant Klebsiella pneumoniae OXA-48 Under Different Growth Conditions

Majid Ahmed AL-Bayati*![]() | Mohammed Fadhil Aboksour
| Mohammed Fadhil Aboksour![]() | Firas Nabeeh Jaafar
| Firas Nabeeh Jaafar![]()
© 2024 The authors. This article is published by IIETA and is licensed under the CC BY 4.0 license (http://creativecommons.org/licenses/by/4.0/).
OPEN ACCESS
Understanding bacterial risk factors, such as biofilm formation and antibiotic resistance, is crucial for treatment. This study investigates biofilm development and gene expression in carbapenem-resistant Klebsiella pneumoniae OXA-48 to understand molecular mechanisms. Two strains of Klebsiella pneumoniae were used in the current study: an environmental strain collected from water samples of army canal in Baghdad, Iraq, identified as a non-resistant strain of K. pneumoniae, and a carbapenem-resistant K. pneumonia OXA-48 (NCTC-13442), which was obtained from the Public Health England culture collection (PHE). Using a tissue culture plate test, biofilms were evaluated under various growth conditions at four different stages of biofilm development: initial attachment (6 hours), microcolony formation (12 hours), maturity (24 hours), and dispersion (48 hours). Genes linked to biofilm development, quorum sensing, and antibiotic resistance were examined for their expression simultaneously using quantitative real-time PCR (qPCR). A significant relation between both biofilm formation phases and carbapenem- resistant Enterobacterales, with the highest biofilm production occurring during the first 24 hours of the maturation stage, also biofilm formation under the studied conditions has a major impact on the diversity of biofilm gene expression quantification studies. The results of the qPCR studies show a correlation between the expression of genes linked to biofilm development and the formation of phenotypic biofilms. These findings show regulation of these genes in significant carbapenem- resistant Enterobacterales pathogens. Also the results showed that the K. pneumoniae OXA-48 CRE strain has a high biofilm formation rate and that genes related to biofilm development are up-regulated during biofilm growth. The findings suggest that the biofilms of this strain successfully adapt to various environmental conditions and play a pathogenic role, linked to multidrug resistance and the successful dissemination of carbapenem-resistant Enterobacterales in common infections.
biofilm, carbapenem, Enterobacteriaceae, K. pneumonia OXA-48, qPCR
The most prevalent and dangerous lower respiratory, bloodstream, and urinary tract infections in both community and healthcare settings are caused by Enterobacterales, which include Klebsiella pneumonia [1].
Enterobacterales have a vicious cycle of resistance development, especially with Klebsiella pneumonia OXA-48. Consumption of carbapenem has increased as a result of increased resistance to all penicillin and cephalosporin antibiotics brought on by the global expansion of extended-spectrum beta-lactamases (ESBLs) among Enterobacterales [2]. As a last- resort antibiotic, carbapenem is commonly used to treat infections brought on by Enterobacterales that are resistant to multiple drugs [3]. Nonetheless, enhanced selection pressure and the global expansion of carbapenem- resistant Enterobacterales (CRE) have resulted from escalating carbapenem intake. Due to their high mortality rates and limited treatment options, CRE infections pose a serious threat to public health. A previous study involving 2462 patients showed that the mortality rate for Klebsiella pneumoniae infections jumped from 21.16% to 42.14% when the bacteria were resistant to carbapenem, compared to carbapenem-susceptible K. pneumoniae [4]. Furthermore, the formation of biofilms is considered by many previous studies as an essential factor for carbapenem-resistant Klebsiella pneumoniaeto colonize in nosocomial infections linked to elevated rates of morbidity and death, especially in cancer patients [1, 4].
Three main categories of carbapenem-resistant (CR) determinants are described by the research that is already available. Class D comprises oxacillinases (OXA)-type enzymes such as OXA-48-like carbapenemases, whereas Class A comprises Klebsiella pneumoniae carbapenemase (KPC), Class B comprises metallo-β-lactamases (MBLs) such as New Delhi MBL (NDM), Verona integrin-encoded MBL (VIM), and imipenemase (IMP) [4, 5]. Genes encoding these CR determinants are mainly associated with mobile genetic elements, which facilitates their quick spread [4, 5]. Several adhesins, hemolysin synthesis, serum resistance, and biofilm formation are examples of CRE virulence factors [3]. An ordered colony of bacteria that forms a matrix and adheres to a surface, either inert or living, is called a biofilm. It is well recognized that bacterial biofilms, which account for 65–80% of all bacterial infections, are a primary cause of numerous illnesses [6]. Generally, many recent studies related to the biofilm formation capabilities of Klebsiella pneumoniae and its high mortality rates due to their implications for treatment resistance [3].
Research has demonstrated that distinct physiological growth conditions and genetic interactions inside biofilms lead to a sharp rise in antimicrobial intolerance and a connection between biofilm formation and antibiotic resistance [7]. While numerous studies have shown that different growth conditions affect the production of biofilms and, consequently, the connection with antibiotic resistance [7], it is unclear how growing circumstances affect the expression of genes relevant to biofilms in CRE. In particular, how the growth environment impacts the expression of the genes in charge of the CRE bacterial cell communication pathways. One kind of communication between bacteria's cells is called quorum sensing. Quorum sensing (QS) is a chemical signaling system that controls bacterial pathogenicity, including motility, adhesion, and biofilm formation and influences gene expression in response to cell density [8].
The autoinducer-2 (AI-2) system, which designates the luxS gene products as AI-2 molecules, is a component of the QS system in Gram-negative bacteria. There are genes in E. coli that are associated with the AI-2 system, and it has been shown that virulence gene expression and biofilm reduction occur when AI-2 QS systems are inhibited [9]. Moreover, it has been shown that in low-nutrient environments, luxS produces AI-2 molecules, which can be exported and tell other cells to form a biofilm. Therefore, it is essential to comprehend how gene expression profiles and biofilm creation are influenced by physiological and growth factors, as well as how these traits are connected to drug-resistant bacteria that are associated with biofilms. Because of the associated poor clinical consequences, the increasing incidence of CRE constitutes a serious risk; outbreaks of CRE have already been reported worldwide [10].
Generally, many previous studies have recorded the difficulty of treating infections caused by biofilm-forming CRKP [3].
The goal of this work is to gain a better understanding of the relationship between biofilm formation and increased resistance in carbapenem-resistant K. pneumoniae isolates, as well as the genes that regulate biofilm-related and antibiotic resistance under different growth conditions.
2.1 Bacterial strains
Two strain of Klebsiellapneumonia were sued in the current study; Carbapenem-resistant K. pneumonia OXA-48 (NCTC-13442) which was collected from the Public Health England culture collection (PHE), and non-resistant K. pneumonia which was isolated from water samples of army canal in Baghdad- Iraq. Both bacterial isolates were cultured on both nutrient agar and MacConkey media.These two isolates were used as positive and negative controls respectively.
2.2 Detection of biofilms formation
The tissue culture plate assay was used to evaluate biofilms formation of K. pneumoniae with some modifications [8, 11]. Bacterial strains were grown for the entire night at 37℃ in 5 ml of nutrient media. 200 µl of inoculation cell suspension was added to four tissue culture plates, and the plates were incubated at 37℃ for varied amounts of time at both static and rocking settings (70 rpm), and there are 6, 12, 24, and 48-hour intervals between each. AB media (which contains AB salts, AB buffers and glucose) and LB media (which contains Tryptone, Salt, and yeast extract) were used in the current study. The broth was taken out of the plate wells after the designated incubation period. To get rid of any non-adherent planktonic cells, the wells were then rinsed twice with sterile phosphate-buffered saline (PBS). Each well was filled with 125 µl of 1% crystal violet and allowed to incubate at room temperature to stain the biofilms. After completely rinsing the stain with sterile water, the dye was re-solubilized with 33% v/v glacial and left to rest at room temperature for half an hour. Eventually, the amount of biofilm produced was ascertained by measuring the optical density of every well at 595 nm optical density. Every test has a negative control that is made up of only media without bacterial suspention. Biofilms were classified as adhering firmly (OD595 > 0.4), moderately (OD595 0.2-0.4), weakly (OD595 < 0.2), or without adhering at all (OD595 = 0). Triplicates of the final values were used to represent them [11].
2.3 RNA isolation and cDNA synthesis
Six milliliters aliquots of the bacterial strains were added to six well of tissue culture plates and incubated for 6, 12, 24, and 48 hours, the plate was statically incubated at 37℃ with a lid sealed. The wells were carefully cleaned with sterile PBS buffer after incubation. RNA isolation (QIAGEN, Germany) provided 100 μL of RNase-free water in the bacterial pellets that were suspended. Following the manufacturer's recommendations, RNA isolation was performed using an RNeasy® Power Biofilm Kit (QIAGEN, Germany) and Turbo DNase was used to decontaminate DNA. Nanodrop spectrophotometer (Thermo Fisher Scientific, USA) was used to detect the absorbance at 260–280 nm, and agarose gel electrophoresis was used to assess the purity of the RNA. Revert Aid H Minus First Stand cDNA Synthesis (Thermo Fisher Scientific, USA) was used to reverse transcribe the RNA to cDNA by the manufacturer's instructions [7, 12, 13].
2.4 Primers of qPCR
The NCBI database (www.ncbi.nim.nih.gov), was used to choose gene-specific primers, and Integrated DNA Technologies-USA (IDT)was the source of primers that were made utilizing the website tools (www.idtdna.com), the primers were selected carefully according to previous studies [4, 11]. Table 1 displays the primers of K. pneumoniae. Using post-PCR agarose gel electrophoresis of the PCR products and melting curve analysis, annealing temperatures for each primer pair were tuned. To determine primer efficiencies, cDNA dilutions of 100, 50, 10, 1 ng/µl, and 0.1 ng/µl were employed in subsequent gene expression calculations [11].
Table 1. K. pneumoniae primer sequences
|
Gene |
Nucleated Sequence (5´- 3´) |
Amplicon Size (bp) |
|
16S rRNA |
F: GGAGAAGGTGGGATGACG |
158 |
|
R: TCCAATCCGGACTACGACGC |
||
|
recA |
F: TCTTCGTCCCGCACCAGTTT |
138 |
|
R: CTTTGACGTTGGTCTGGCGG |
||
|
tonB |
F: TTCCCACCGGAAGCGAAAGT |
101 |
|
R: TTCCGAAGTTCAGCCCGCTA |
||
|
blaOXA- 48 |
F: GCTTGATCGCCCTCGATT |
122 |
|
R: GATTTGCTCCGTGGCCGAAA |
||
|
aph |
F: GGTGGTATTCGCATTTCGGC |
158 |
|
R: GAGTATCCCGTTGTAGCCCG |
||
|
ST- 48 |
F: CTGACCGTCACGGGTAACAA |
74 |
|
R: TTGATGTGGCGGTATGCAGT |
||
|
aac |
F: GTTCAAACAGGCACACCGAC |
114 |
|
R: AGCTGTTCGGCTACGACAAA |
||
|
qnrD |
F: GACTATCCGTTCTGAGGCGG |
107 |
|
R: TGCGTAAAAGCACAACCGTG |
2.5 Quantitative Real-time PCR (qPCR)
By using qPCR, the transcript levels of the eight genes in K. pneumoniae were assessed in AB and LB media at four different growth phases, namely after 6, 12, 24, and 48 hours of incubation. The genes for biofilm formation, quorum sensing, stress survival, and antibiotic resistance were categorized for each strain. Sensi FAST SYBR NO-ROX Kit (BioLine, USA) was used for the qPCR reaction [13]. Each sample had a reaction volume of 20 μl, 200 ng/ml of cDNA, and 400 nM of each primer separately. In comparison to the growth shown at 12, 24, and 48 hours, the calibrator sample at 6 hours had the least amount of biofilm. On 96-well plates, each reaction was run in technical triplicate with two biological duplicates. The mean of each triple was used to express the final results.
2.6 Statistical analysis
IBM SPSS version 22.0 (SPSS Inc., Chicago, IL, USA) was used to enter and analyze the outcome data. In regard to frequency, proportions, means, as well as standard deviations, descriptive statistics were employed. The Chi-square test was performed to assess the associations between the existence of resistance-gene-carrying samples and the general and clinical features of the patients [11].
3.1 Biofilm quantitative analysis
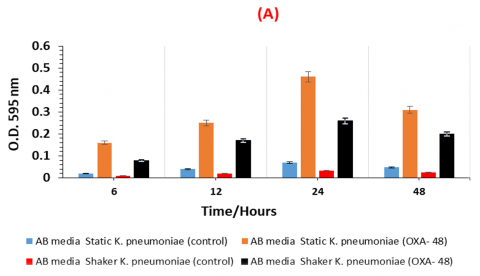
The quantities of biofilm produced by non-resistant K. pneumoniae and Carbapenem-resistant K. pneumonia OXA-48 in AB and LB media are displayed in Figure 1. It is clear that the K. pneumoniae OXA-48 strain exhibited the highest turbidity at all the chosen times (6, 12, 24, and 48 hours) compared to the non-resistant K. pneumoniae strain.Moreover, static method was mentioned as a best method comparing with shaker method as a result of the current results, while non-resistant K. pneumoniae didn't record any significant value under 595 nm. Table 2 illustrates the mean values of biofilm formation of the both strains.
Figure 1. Biofilm formation of K. pneumoniae strains under shaking and static conditions in both nutrient-poor AB medium (A) and nutrient-rich LB media (B)
Table 2. Biofilm formation means of carbapenem resistant and non- resistant K. pneumoniae in AB and LB media
|
Media |
Time (hours) |
Static |
Shaker |
||
|
Carbapenem resistant strain |
Non-Carbapenem resistant strain |
Carbapenem resistant strain |
Non-Carbapenem resistant strain |
||
|
AB |
6 |
0.16 ± 0.01 |
0.02 ± 0.001 |
0.08 ± 0.01 |
0.01 ± 0.001 |
|
12 |
0.25 ± 0.025 |
0.04 ± 0.002 |
0.17 ± 0.005 |
0.02 ± 0.002 |
|
|
24 |
0.46 ± 0.02* |
0.07 ± 0.001* |
0.26 ± 0.01* |
0.03 ± 0.002 |
|
|
48 |
0.31 ± 0.01 |
0.048 ± 0.005 |
0.2 ± 0.0015 |
0.026 ± 0.003 |
|
|
LB |
6 |
0.38 ± 0.015 |
0.05 ± 0.005 |
0.2 ± 0.01 |
0.03 ± 0.002 |
|
12 |
0.46 ± 0.01 |
0.09 ± 0.001 |
0.35 ± 0.015 |
0.05 ± 0.002 |
|
|
24 |
0.73 ± 0.026* |
0.12 ± 0.001* |
0.51 ± 0.016* |
0.09 ± 0.001* |
|
|
48 |
0.58 ± 0.03 |
0.09 ± 0.002 |
0.4 ± 0.02 |
0.07 ± 0.1 |
|
Note: * means significant in P value 0.01.
Table 2 demonstrates that K. pneumoniae OXA-48 attached strongly (OD595 nm > 0.75) in all investigated scenarios, but the non-resistant strain adhered weakly (OD595 nm < 0.15). Depending on their growth stages, the K. pneumoniae OXA-48 strain displayed varying degrees of biofilm formation. In comparison to the previous time points of 6, 12, and 48 hours, the target sample at 24 hours is statistically significant. Furthermore, 24 hours treatments were mentioned as significant values in all the treatment conditions. As well as, the strongest biofilm mass of K. pneumonia OXA-48 were recognized after 24 hours in both AB and LB media as 0.48 and 0.75 respectively.
3.2 Stability of RNA, selection of reference genes, primer efficiencies, specificity of primers for qPCR
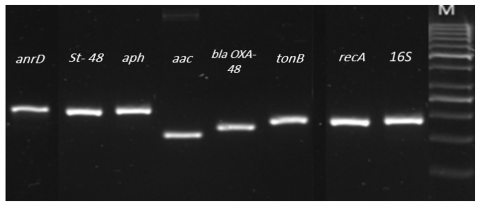
RNA of sufficient quality and quantity was generated under various development circumstances and time intervals. Among the chosen candidates, the most stable reference (housekeeping) genes were found using the geNorm technique. According to previously study [14], a gene expression normalization factor was calculated for each target gene using the geometric mean of a predetermined number of genes. As reference candidates, the reference genes for K. pneumoniae OXA-48, 16SRNA, recA, and tonB were chosen. GeNorm indicated that the most consistently expressed genes in all samples were 16S RNA and recA, as they had the lowest combined M-values (0.21). K. pneumoniae OXA-48 gene primer efficiency was evaluated by dilution of cDNA at increasing concentrations (100 ng/L). Using melting curve analysis following qPCR amplification, primer sets for target genes in K. pneumonia OXA-48 were evaluated for specificity. Each tested pair of primers yielded a single peak and product due to strain. All primer pairs amplified a single target gene product according to the endpoint qPCR findings obtained by agarose gel electrophoresis, see Figure 2.
Figure 2. PCR results of the carbapenem resistant genes of K. pneumoniae OXA-48
Note: lane M is DNA ladder.
3.3 Expression levels of antibiotic resistant and biofilm associated genes in K. pneumoniae OXA-48
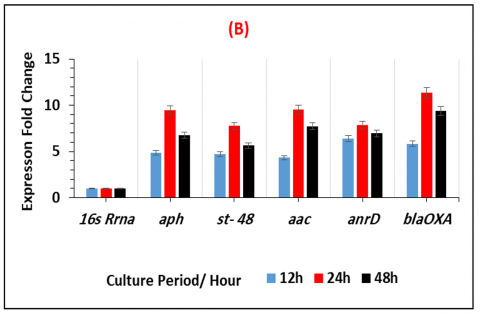
The fold expression values for K. pneumonia OXA- 48 show the expression levels of chosen genes at 12 h, 24 h, and 48 h of biofilm formation as well as in two growth media. K. pneumoniae OXA-48 shows varying amounts of gene expression over time in Figure 3. The early adherence phase (12 h) was when upregulation was first observed, increasing at the mid-adherence phase (24 h), and then somewhat declining during the biofilm dispersion phase (48 h). All growth media and gene categories including biofilms, and quorum sensing showed this pattern of gene regulation. After a day, quorum sensing genes, including anrD and aac, were markedly upregulated in both media. At 24 hours, the antibiotic resistance gene blaOXA-48displayed the highest level of overexpression. When K. pneumoniae OXA-48 was grown in the nutrient-rich LB medium as opposed to the nutrient-poor AB medium, all gene categories showed significant increases (see Figure 3 and Table 3).
Figure 3. The expression fold change of the selected genes related to biofilm development, quorum sensing, and antibiotic resistance in K. pneumoniae OXA-48 grown on (A) nutrient-rich media (LB) and (B) nutrient-poor media (AB)
Table 3. Fold change in levels of K. pneumonia OXA-48 biofilm genes as a function to the elected genes in both LB and AB media
|
Genes |
|
AB Media |
LB Media |
||||
|
Type |
12 hrs |
24 hrs |
48 hrs |
12 hrs |
24 hrs |
48 hrs |
|
|
16S |
REF |
1 |
1 |
1 |
1 |
1 |
1 |
|
aph |
TRG |
2.83±0.01 |
7.71±0.019* |
3.88±0.032** |
4.88±0.03 |
9.48±0.01* |
6.75±0.015** |
|
St-48 |
TRG |
3.35±0.019 |
5.21±0.012* |
4.22±0.002** |
4.7±0.02 |
7.75±0.03* |
5.65±0.011** |
|
aac |
TRG |
2.19±0.06 |
7.33±0.022* |
4.31±0.002** |
4.32±0.013 |
9.54±0.012* |
7.74±0.021** |
|
anrD |
TRG |
3.34±0.022 |
5.19±0.042* |
4.04±0.003** |
6.37±0.032 |
7.85±0.003* |
6.95±0.012** |
|
Bla OXA-48 |
TRG |
3.93±0.022 |
9.78±0.002* |
7.23±0.06** |
5.82±0.012 |
11.35±0.02* |
9.41±0.013** |
Klebsiella pneumoniae and other Enterobacterales frequently form biofilms [11]. Biofilm growth encourages survival. Hospital environments and internal patients are more susceptible to bacterial infections [15]. Because they are common in both community-acquired infections and hospital-acquired infections, multidrug-resistant CRE, like K. pneumoniae OXA-48, poses a serious health risk. Global epidemics of CRE have been seen in the past [16]. It was possible to accurately document the clinical risk factors for bacterial infections linked to CRE; however, bacterial risk variables such as antibiotic susceptibility, resistance mechanisms, and biofilm formation were not [16]. Numerous clinical illnesses have been linked to biofilms, and there is increasing evidence that biofilms play a role in pathogenesis, particularly in chronic infections [17]. There aren't many CRE infections. To treat biofilm-associated illnesses brought on by CRE pathogens, which have poor clinical outcomes and few available treatment options, it is essential to comprehend bacterial characteristics, especially biofilms. According to our research, K. pneumoniae OXA-48, the resistant gene blaOXA-48, developed robust biofilms and there may be a connection between biofilm formation and antimicrobial resistance genes [15]. Comparing genes to non-resistant control strains was done. A similar tendency of enhanced biofilm development has been observed in previous research on multidrug-resistant Enterobacteriaceae [17]. K. pneumoniae OXA-48 demonstrated the strongest biofilm growth, and at 24 hours, the resistance gene blaOXA-48 displayed the highest overexpression. Resistance genes have been demonstrated to affect the formation of biofilms, which could account for this phenomenon. In this work, test bacteria were incubated under both shaking and static conditions to investigate the effects of aeration and nutrient distribution on biofilm formation [15].
The current study showed that when the CRE strain was cultured, it produced large biofilms. This feature may be explained by the fact that facultative anaerobes prefer static incubation [11]. According to the biofilm development stages of attachment (6 hours), exopolysaccharide production (12 hours), and microcolony formation, the CRE strain's biofilm analysis revealed varying degrees of biofilms. At the 48-hour point, the biofilm began to detach and disperse after 24 hours of maturation and stabilization of a three-dimensional structure. The formation of the biofilm matched previous studies on Gram-negative bacteria [18].
Moreover, the highest strain was observed in CRE. The amount of biofilm seen in drug-resistant E. coli isolates [10] and multidrug-resistant P. aeruginosa isolates [17, 19] at the biofilm maturation stage (24 hours of incubation) is comparable. During the 24-hour biofilm maturation period, K. pneumoniae OXA-48 up-regulates the biofilm-related genes aph and st-48, suggesting their role in biofilm formation [20].
An essential small cytoplasmic protein is encoded by the aph gene. Controls the growth of biofilms by secreting signals. Fimbriae, a crucial virulence factor for attachment, adhesion, colonization, and biofilm formation, are encoded by St-48 [20].
Similar to recent work on a drug-resistant K. pneumoniae clinical isolate, QS genes aaC and anrD were shown to be substantially elevated during biofilm maturation (24 h in all growth conditions studied) in K. pneumoniae OXA-48. Moreover, all the selected genes that were used in the current study were recorded as the most common and popular genes in the previous studies which were dealing with the same object [11, 20, 21]. Gene expression is dependent on nutritional availability and environmental conditions and governs biofilm formation, polysaccharide synthesis, and metabolic pathways [21]. The strain's hypervirulence and resistance during biofilm development, which lead to efficient dispersion in various environments common infections may be the cause of its high-level overexpression of blaOXA, which encodes for carbapenem resistance [22].
The carbapenem-resistant blaOXA gene is most overexpressed in OXA-48 when the biofilm develops and grows for 24 hours. Hypervirulence may be associated with blaOXA-48overexpression at high levels. This strain process displays resistance in the biofilm generation phase. These strains differ in the antibiotic-resistant genes they carry because of their multidrug resistance. Their plasmids include resistant determinants that readily combine with other bacteria to produce widespread antibiotic resistance [22]. The blaOXA gene is usually adopted by conjugative plasmids, leading to multiple determinants and multidrug resistance [23]. The QS gene (aac and anrD) and its expression during the biofilm formation stages, biofilm maturation genes (aph and st-48), and antibiotic resistance genes blaOXA-48were the main topics of this work. Our investigation is limited by the biofilm formation on two resistant bacteria, which is restricted to a specific cluster of genes.
According to our research, several genes might be involved in the production of biofilms. To better understand the physiological impact of biofilms and distinguish gene expression, more study is required. One example of this would be to correlate the magnitude of the biofilm with the resistance determinants in CRE.
This could shed light on the role that resistance genes play in the production of biofilms. This has the potential to advance our knowledge of the molecular mechanism behind the long-term formation of biofilms. According to our research, the stages of biofilm development are linked to the CRE strain's gene expression. Across all growth conditions examined, gene clusters were primarily elevated during biofilm formation (24 h) and downregulated during detachment (48 h). The study demonstrates how gene expression profiles and morphological changes can be correlated using qPCR.
The static method was mentioned more suitable in the biofilm formation of bacterial isolates comparing with shaker method. The variety of biofilm gene expression quantification studies is significantly impacted by biofilm formation under the studied conditions, and there is a significant correlation between both biofilm formation phases and carbapenem-resistant Enterobacterales, with the highest biofilm.Also, the expression of a group of genes linked to the formation of biofilms in enteric K. pneumoniae OXA strains of carbapenem-resistant Gram-negative bacteria was examined. In K. pneumoniae OXA-48 strains, biofilm production is accompanied by a rise in biofilm formation genes. The most important element influencing the variability of biofilm gene expression quantification assays, according to our research, is biofilm formation. Biofilm growth and the expression of particular genes related to biofilm, quorum sensing, and antibiotic resistance are correlated, according to qPCR research.
This morphological analysis can demonstrate differences in gene regulation in significant CRE pathogens, as well as changes in gene expression during biofilm formation. Our findings, supported by the literature we have reviewed, suggest thatK. pneumoniae OXA-48, which forms biofilms effectively under various growth conditions, may develop antibiotic resistance. This increases the likelihood that CRE pathogens will successfully spread and acquire multidrug resistance. The results also show that multidrug resistance and the effective dissemination of carbapenem-resistant Enterobacterales in common infections are linked to the biofilms of this strain, which efficiently adapt to various environmental conditions and play a pathogenic role.
The author would like to thank Mustansiriyah University (www.uomustansiriyah.edu.iq) Baghdad-Iraq for its support in the present work and the Ministry of Higher Education and Scientific Research in Iraq/Baghdad.
[1] Magiorakos, A.P., Burns, K., Baño, J.R., Borg, M., Daikos, G., Dumpis, U., Lucet, J.C., Moro, M.L., Tacconelli, E., Simonsen, G.S., Szilágyi, E., Voss, A., Weber,J.T. (2017). Infection prevention and control measures and tools for the prevention of entry of carbapenem-resistant Enterobacteriaceae into healthcare settings: Guidance from the European Centre for Disease Prevention and Control. Antimicrobial Resistance & Infection Control, 6(1): 113. https://doi.org/10.1186/s13756-017-0259-z
[2] Badr, A.A., Abbas, T., AboKsour, M.F. (2022). Diagnosing bacteria samples using data mining: Review study. In 2022 5th International Conference on Engineering Technology and Its Applications (IICETA), Al-Najaf, Iraq, pp. 58-63. https://doi.org/10.1109/IICETA54559.2022.9888705
[3] Li, L., Gao, X., Li, M., Liu, Y., Ma, J., Wang, X., Yu, Z., Cheng, W., Zhang, W., Sun, H., Song, X., Wang, Z. (2024). Relationship between biofilm formation and antibiotic resistance of Klebsiella pneumoniae and updates on antibiofilm therapeutic strategies. Frontiers in Cellular and Infection Microbiology, 14: 1324895. https://doi.org/10.3389/fcimb.2024.1324895
[4] Xu, L., Sun, X., Ma, X. (2017). Systematic review and meta-analysis of mortality of patients infected with carbapenem-resistant Klebsiella pneumoniae. Annals of Clinical Microbiology and Antimicrobials, 16(1): 18. https://doi.org/10.1186/s12941-017-0191-3
[5] Michaelis, C., Grohmann, E. (2023). Horizontal gene transfer of antibiotic resistance genes in biofilms. Antibiotics, 12(2): 328. https://doi.org/10.3390/antibiotics12020328
[6] AboKsour, M.F., Al-Marjani, M.F., Rheima, A.M. (2024). Preparation and effects of manganese oxide nanoparticles against quinolone-resistant bacteria isolated from hospital wastewater. Al-Rafidain Journal of Medical Sciences, 6(2): 94-100. https://doi.org/10.54133/ajms.v6i2.728
[7] Al-Bayati, M., Samarasinghe, S. (2022). Biofilm and gene expression characteristics of the carbapenem-resistant enterobacterales, Escherichia coli IMP, and Klebsiella pneumoniae NDM-1 associated with common bacterial infections. International Journal of Environmental Research and Public Health, 19(8): 4788. https://doi.org/10.3390/ijerph19084788
[8] Juszczuk-Kubiak, E. (2024). Molecular aspects of the functioning of pathogenic bacteria biofilm based on quorum sensing (QS) signal-response system and innovative non-antibiotic strategies for their elimination. International Journal of Molecular Sciences, 25(5): 2655. https://doi.org/10.3390/ijms25052655
[9] Mirghani, R., Saba, T., Khaliq, H., Mitchell, J., Do, L., Chambi, L., Diaz, K., Kennedy, T., Alkassab, K., Huynh, T., Elmi, M., Martinez, J., Sawan, S., Rijal, G. (2022). Biofilms: Formation, drug resistance and alternatives to conventional approaches. AIMS Microbiology, 8(3): 239-277. https://doi.org/10.3934/microbiol.2022019
[10] Bocanegra-Ibarias, P., Garza-González, E., Morfín-Otero, R., Barrios, H., Villarreal-Treviño, L., Rodríguez-Noriega, E., Garza-Ramos, U., Petersen-Morfin, S., Silva-Sanchez, J. (2017). Molecular and microbiological report of a hospital outbreak of NDM-1-carrying Enterobacteriaceae in Mexico. PLOS One, 12(6): e0179651. https://doi.org/10.1371/journal.pone.0179651
[11] Samarasinghe, S., Reid, R., Al-Bayati, M. (2019). The anti-virulence effect of cranberry active compound proanthocyanins (PACs) on expression of genes in the third-generation cephalosporin-resistant Escherichia coli CTX-M-15 associated with urinary tract infection. Antimicrobial Resistance & Infection Control, 8: 181. https://doi.org/10.1186/s13756-019-0637-9
[12] Rahdar, H.A., Malekabad, E.S., Dadashi, A.R., Keikha, M., Kazemian, H., Karami-Zarandi, M. (2019). Correlation between biofilm formation and carbapenem resistance among clinical isolates of Klebsiella pneumoniae. Ethiopian Journal of Health Sciences, 29(6): 711-718. https://doi.org/10.4314/ejhs.v29i6.11
[13] Maeyama, R., Mizunoe, Y., Anderson, J.M., Tanaka, M., Matsuda, T. (2004). Confocal imaging of biofilm formation process using fluoroprobedEscherichia coli and fluoro‐stained exopolysaccharide. Journal of Biomedical Materials Research Part A, 70A(2): 274-282. https://doi.org/10.1002/jbm.a.30077
[14] Vandesompele, J., De Preter, K., Pattyn, F., Poppe, B., Van Roy, N., De Paepe, A., Speleman, F. (2002). Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biology, 3(7). https://doi.org/10.1186/gb-2002-3-7-research0034
[15] Lebeaux, D., Ghigo, J.M., Beloin, C. (2014). Biofilm-related infections: Bridging the gap between clinical management and fundamental aspects of recalcitrance toward antibiotics. Microbiology and Molecular Biology Reviews, 78(3): 510-543. https://doi.org/10.1128/mmbr.00013-14
[16] Hussein, K., Rabino, G., Eluk, O., Warman, S., Reisner, S., Geffen, Y., Halif, L., Paul, M. (2017). The association between infection control interventions and carbapenem-resistant Enterobacteriaceae incidence in an endemic hospital. Journal of Hospital Infection, 97(3): 218-225. https://doi.org/10.1016/j.jhin.2017.07.018
[17] Vestby, L.K., Grønseth, T., Simm, R., Nesse, L.L. (2020). Bacterial biofilm and its role in the pathogenesis of disease. Antibiotics, 9(2): 59. https://doi.org/10.3390/antibiotics9020059
[18] Anderson, G.G., Palermo, J.J., Schilling, J.D., Roth, R., Heuser, J., Hultgren, S.J. (2003). Intracellular bacterial biofilm-like pods in urinary tract infections. Science, 301(5629): 105-107. https://doi.org/10.1126/science.1084550
[19] Musafer, K., Jaafar, N., Al-Bayati, M. (2022). Association of biofilm inducer with blaVIM, blaIMP, and blaNDM in Pseudomonas aeruginosa isolates. Archives of Razi Institute, 77(5): 1723-1728. https://doi.org/10.22092/ari.2022.358104.2153
[20] Gharrah, M.M., El-Mahdy, A.M., Barwa, R.F. (2017). Association between virulence factors and extended spectrum beta-lactamase producing Klebsiella pneumoniae compared to nonproducing isolates. Interdisciplinary Perspectives on Infectious Diseases. https://doi.org/10.1155/2017/7279830
[21] Chen, L., Wilksch, J.J., Liu, H., Zhang, X., Torres, V.V., Bi, W., Mandela, E., Cao, J., Li, J., Lithgow, T., Zhou, T. (2020). Investigation of LuxS-mediated quorum sensing in Klebsiella pneumoniae. Journal of Medical Microbiology, 69(3): 402-413. https://doi.org/10.1099/jmm.0.001148
[22] Yong, D., Toleman, M.A., Giske, C.G., Cho, H.S., Sundman, K., Lee, K., Walsh, T.R. (2009). Characterization of a new metallo-β-lactamase gene, blaNDM-1, and a novel erythromycin esterase gene carried on a unique genetic structure in Klebsiella pneumoniae sequence type 14 from India. Antimicrobial Agents and Chemotherapy, 53(12): 5046-5054. https://doi.org/10.1128/AAC.00774-09
[23] Zhou, Y., Smith, D.R., Hufnagel, D.A., Chapman, M.R. (2013). Experimental manipulation of the microbial functional amyloid called curli. In: Bacterial Cell Surfaces: Methods and Protocols, pp. 53-75. https://doi.org/10.1007/978-1-62703-245-2_4