Nonlinear Height-Diameter Models for Anthocephalus macrophyllus in Indonesia

Yonky Indrajaya*![]() | Mohamad Siarudin
| Mohamad Siarudin![]() | Muhammad Abdul Qirom
| Muhammad Abdul Qirom![]() | Arif Nirsatmanto
| Arif Nirsatmanto![]() | Sri Sunarti
| Sri Sunarti![]() | Noor Khomsah Kartikawati
| Noor Khomsah Kartikawati![]() | Anto Rimbawanto
| Anto Rimbawanto![]() | Liliek Haryjanto
| Liliek Haryjanto![]() | Toni Herawan
| Toni Herawan![]() | Asri Insiana Putri
| Asri Insiana Putri![]() | Fajar Lestari
| Fajar Lestari![]() | Wawan Halwany
| Wawan Halwany![]() | Pamungkas Buana Putra
| Pamungkas Buana Putra![]() | Dian Pratiwi
| Dian Pratiwi![]()
© 2024 The authors. This article is published by IIETA and is licensed under the CC BY 4.0 license (http://creativecommons.org/licenses/by/4.0/).
OPEN ACCESS
Plantation of Anthocephalus macrophyllus (Roxb.) Havil. is relatively extensive in Indonesia. Reliable and updated height–diameter models are required to quantify the growth of A. macrophyllus. In this study, we developed the tree height-diameter at breast height (DBH) models for A. macrophyllus in Indonesia to better understand the developmental dynamics and management using twenty-seven models. A total of 2961 total height and DBH measurements were used and randomly split into two datasets. Eighty percent of the measurements were utilized for the initial parameter estimation of the twenty-seven models, while the remaining 20% were used for model validation. The quality of the models was assessed using the following metrics: Root Mean Square Error (RMSE), Mean Absolute Percentage Error (MAPE), coefficient of determination (R2), adjusted coefficient of determination (R2-adj), Akaike Information Criterion (AIC), and Bayesian Information Criterion (BIC). The Mean Difference (MD) for each DBH class (5 cm interval) and the overall MD were calculated for model validation. The results of this research showed that three-parameter models of height-diameter metrics are slightly better than two-parameter models. The best model is the Chapman-Richards (R2: 0.937; MAPE: 9.98; RMSE: 1.1) and Huang model (R2: 0.937; MAPE: 9.96; RMSE: 1.1), which is reliable and has been applied in most of the three stands in the literature. Since correct height-diameter models can increase the efficiency of forest inventories, improve growth forecasts, and help to build sustainable management methods, these results have important consequences for forest management and policy-making. Moreover, the acceptance of these models can guide policy decisions on long-term planning for the preservation and use of A. macrophyllus plantations, resource allocation, and reforestation initiatives.
Anthocephalus macrophyllus (Roxb.) Havil., height-diameter models, tree growth models, Chapman-Richards model
Anthocephalus macrophyllus (Roxb.) Havil. commonly referred to as red jabon, is one of the species of the genus Anthocepalus, which originally spread in Sulawesi, Mollucas, and Papua [1]. Another species from the genus Anthocephalus, Anthocephalus cadamba, or commonly white jabon, is an indigenous tropical tree species found in South and Southeast Asia, including most parts of Indonesia [2, 3]. Although the two species can be regarded as synonymous, foresters and farmers can clearly see variations in appearance and growth [4]. This genus has been intentionally grown in Java, Kalimantan, Sumatera, Sulawesi, Sumbawa, and Papua [5]. Jabon is a tree species with great potential for use in industrial forest plantations and reforestation in Indonesia. This is due to its fast growth, ability to thrive in different environments, ease of silvicultural treatment, and resistance to major pests and diseases [3]. The wood of both jabon species is also reported as a potential raw material for fiber wood industries, such as pulp and paper [6, 7]. In the future, the species is anticipated to gain greater significance in the timber sector, particularly as the availability of raw materials for woodworking from natural forests is projected to diminish. Extensive jabon plantations are located in the provinces of North Sumatra, Riau, and Central Kalimantan. Presently, jabon is extensively cultivated by farmers, particularly in Kalimantan and Java. Nair [8] reported that jabon is commonly cultivated in certain regions of Java as a substitute for teak trees that exhibit limited growth after being harvested.The red jabon is a large tree with a broad umbrella-like crown that resembles an umbrella and a cylindrical trunk when examining its physical characteristics. The branches of the tree were arranged in layers. With round and upright trunks reaching 70 - 80% and trunk circumference exceeding more than 150 cm (diameter greater than 50 cm), red jabon trees can reach 40 meters [1]. The light-tolerant pioneer plant red jabon may survive in lowlands up to 50–1000 m above sea level. Red jabon flourished abundantly on karst hills next to hot springs in Hungoyono, Bone Bolango Regency, Gorontalo. Maleo birds (Macrocephalon maleo) nest on these sites [1]. While white jabon typically reaches a moderate size of 15-20cm and possesses a rounded crown. White jabon often reach a maximum height of 45 meters, with a stem diameter of 100 to 160 cm and a little buttress that can be up to 2 meters high [2]. The branches are horizontally elongated, organized in layers, and finish directly at the top. The bark of the tree is characterized by a deep grey color, with extensive cracks and thin scales that peel off.
Previous studies on red jabon in Indonesia have primarily focused on its growth at the seedling stage, examining the effects of various media [9-11], fertilization practices [12], and watering frequency [13]. However, there is a gap in our understanding how these trees perform after planting in the field. The study conducted by A’ida et al. [14] analyzed the progeny tests on the growth of red jabon plants in the field, but it only focused on 2-year-old red jabon and has not yet been able to model the relationship between height and stem diameter. Research on 4-years-old red jabon for carbon sequestration potential [15], and biomass source for bioenergy [16], has similarly lacked modeling of stem growth, which is a major component of biomass and carbon.Additional studies have highlighted other aspects of red jabon. Cahyono et al. [9] investigated the wood quality of red jabon aged 8-10 years, revealing the potential of the species as a future wood supply owing to its straight trunk and long clear bole, attributed to its rapid growth. However, this study did not include growth modeling of tree stems. Another study analyzed the genetic diversity [17] and extinction risk [18] of red jabon, aiming to provide recommendations for conservation strategies for this species.Diameter at breast height (DBH) and tree height (H) are crucial attributes used in forest inventories to estimate stand volume, biomass, carbon, and other related characteristics [19-22]. These variables are crucial prerequisites for the development of forest management plans [23]. Although the determination of DBH may be easily achieved with high precision and at low cost, measuring tree height is more challenging, especially in forests with mixed tree species and in areas with dense canopy cover or uneven terrain [20, 24]. Concerning the importance of red jabon for forest plantation, efforts to improve tree productivity are necessary either based on silviculture or genetics. In terms of genetics, the tree breeding program of red jabon has been initiated by establishing a first-generation breeding seedling orchard [25]. After practicing a series of genetic thinning selections, the orchard can be used as a seed orchard to produce genetically improved seeds for plantation. However, information on the growth model of red jabon, which is important not only for silviculture practices but also for accelerating genetic selection in breeding, is still limited.
This research aims to provide useful insights into the growth dynamics and management techniques of red jabon by examining growth patterns and investigating the correlation between tree diameter and height. By investigating the correlation between DBH and height in red jabon plantations in Indonesia, this study aimed to provide significant insights for optimizing forest management methods and enhancing the understanding of growth dynamics in this commercially important species by examining the development patterns and analyzing the link between tree diameter and height in these stands. The developed pattern could be used to develop an effective breeding strategy for multiple-trait genetic selection and to optimize age-age genetic correlations based on height and diameter growth. Therefore, this inquiry will enhance the existing knowledge on the development patterns of red jabon, assisting forest managers, policymakers, researchers, and tree breeders in developing more efficient strategies for sustainable forestry practices and optimizing the productivity of the stands. This study aims to provide useful insights into the growth dynamics and management techniques of this economically significant species by examining growth patterns and investigating the correlation between tree diameter and height.
Recent research on height-diameter models has shown their crucial role in precisely estimating forest biomass and tree growth. Nevertheless, because they rely on region-specific data and simplified assumptions, current models, such as those established by Peng et al. [26] and Sharma and Parton [22], often show limits when applied across many species and ecological situations. Reduced precision results from many models fail to include fundamental covariates such as stand density and soil properties. Furthermore, it is not practical in dense or inaccessible forests and labor-intensive conventional DBH measuring techniques. By creating more flexible three-parameter models, which offer a more accurate depiction of tree development dynamics, this study seeks to fill these voids. Moreover, this study improves the applicability of height-diameter models for different forest types by using a thorough dataset of red jabon in Indonesia. Combining modern technology and variables, this study aimed to increase model accuracy and offer practical information for policy making and sustainable forest management.
2.1 Research sites and stand characteristics
This research was conducted in a first-generation Breeding Seedling Orchard (BSO) of red jabon located in Wonogiri District, Central Java Province, with geographical coordinates of 7°80′ S, 110°93′ E and an altitude of 141 m above sea level. According to the Schmidt-Fergusson climate classification, it has climate type C with an average annual rainfall of 1,645 mm, and an average daily temperature of 26.5℃. The BSO was established on an area of 1.4 hectares with the vertisol type of soil.
This BSO consisted of 55 family genotypes from the Konawe provenance of South Sulawesi, which was laid out in a randomized completely block design (RCBD), with 4 replications/block, and 4 trees per plot. At the beginning of the plot establishment, planting was done at a spacing of 4 × 2 m. Two series of thinning were carried out simultaneously at 46, and 60 months as first and second, thinning, respectively. The purpose of this stage was to retain one of the best trees within the plot; therefore, thinning was carried out by cutting the poorest phenotype trees within the plot at rates of two out of four trees and one out of two trees of the remaining tree during the first, and second thinning, respectively.
2.2 Data collection
This research was carried out in a red jabon first-generation Breeding Seedling Orchard (BSO) in Wonogiri District, Central Java Province. From trees ranging in age from 6 to 108 months. Maintaining a representative sample across several DBH categories and guaranteeing healthy trees free from significant pests or diseases were the standards for choosing plants. This method guaranteed thorough and consistent data collecting, therefore providing a strong basis for model building and validation.
Data were collected from all remaining trees in the BSO at 6, 12, 24, 36, 52, 60, 72, and 108 months of age. The number of observed trees in each measurement period varied depending on the series of thinning and mortality. Measurement parameters include tree height and diameter at breast height (DBH). Tree height was measured using a pole, while a phi-band meter was used to measure the tree diameter. We collected 2961 paired data points on the height and diameter of the individual trees. To select trees from the available data pairs for model building and validation, we first sorted the sample trees by diameter from smallest to largest. Then, trees with an order of multiples of 5 were selected to be used as data for validation.
2.3 Data analysis
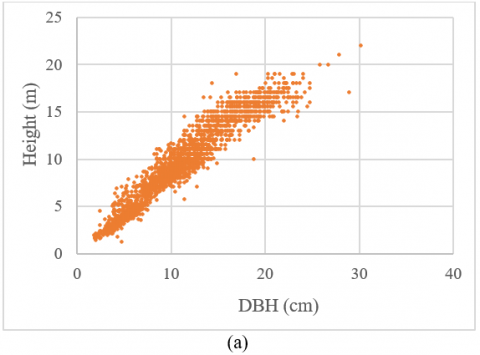
The relationship between tree height and diameter is typically nonlinear. As the tree grows, its growth rate increases from a minimum value, reaching a maximum at an inflection point, and then gradually decreases towards zero as it approaches the upper asymptote . The relationship between the height and DBH of A. macrophyllus was analyzed by plotting the total height against the DBH, following the methods proposed by [26] and Lumbres et al. [28]. A standard sigmoidal concave curve was identified (Figure 1). Twenty-seven candidate models were chosen from prior research based on their suitable mathematical characteristics, potential biological explanation of parameters, and accurate prediction for a tree height-diameter correlation in the literature (Table 1). The chosen models are categorized as follows: Models M1 to M15 are two-parameter models, and models M16 to M27 are three-parameter models.
Seven model performance criteria were utilized to choose models that more accurately depict the correlation between tree heights and diameters (Table 2). Typically, models that have the lowest average values of RMSE, MAPE, AIC, and BIC, and the greatest average values of R2 and R2-adj, are considered to be the best [19, 29]. Following Lumbres et al. [28], we also used MD to validate the models with 20% of the dataset.
Table 1. Selected non-linear height-diameter functions
|
No. |
Model |
Equation |
Source |
|
1 |
Stoffels |
h=1.3+b1DBHb2 |
[30] |
|
2 |
Naslund |
h=1.3+(DBHb1+b2DBH)2 |
[24] |
|
3 |
Huang |
h=1.3+ˉb1DBHb2+DBH |
[31] |
|
4 |
Curtis |
h=1.3+b1(DBH1+DBH)b2 |
[32] |
|
5 |
Curtis |
h=1.3+b1(1+1DBH)D2 |
[32] |
|
6 |
Curtis |
h=1.3+b1DBH(1+DBH)b2 |
[32] |
|
7 |
Meyer |
h=1.3+b1[1−exp(−b2DBH)] |
[33] |
|
8 |
Wykoff |
h=1.3+exp(b1+b21+DBH) |
[34] |
|
9 |
Bates Watts |
h=1.3+b1DBH(1+DBH)+b2DBH |
[35] |
|
10 |
Huang |
h=1.3+b1DBHexp(−b2DBH) |
[31] |
|
11 |
Staudhammer LeMay |
h=1.3+exp(b1+b2DBH) |
[36] |
|
12 |
Exponential |
h=1.3+b1exp(b2DBH) |
[37] |
|
13 |
El Mamoun |
h=1.3+b1[ln(1+DBH)]b2 |
[38] |
|
14 |
El Mamoun |
h=1.3+(b1+b2DBH)−5 |
[38] |
|
15 |
Ogana |
h=1.3+((b1DBH)b2)−1 |
[39] |
|
16 |
Huang |
h=1.3+b11+b2DBH−b3 |
[31] |
|
17 |
Huang |
h=1.3+DBH2b1+b2DBH+b3DBH2 |
[31] |
|
18 |
Huang |
h=1.3+b11+b2exp(−b3DBH) |
[31] |
|
19 |
Weibull |
h=1.3+b1(1−exp(−b2DBHb3)) |
[40] |
|
20 |
Chapman-Richards |
h=1.3+b1(1−exp(−b2DBH))b3 |
[26] |
|
21 |
Titus |
h=1.3+b1exp(−b2exp(−b3DBH)) |
[41] |
|
22 |
Larsen |
h=1.3+exp(b1+b2DBHb3) |
[42] |
|
23 |
Ratkowsky |
h=1.3+exp(b1+b2DBH+b3) |
[19] |
|
24 |
Stage |
h=1.3+b1exp(−b2DBH−b3) |
[43] |
|
25 |
Artroschenko |
h=1.3+b1√DBH+b2DBH+b3DBH2 |
[19] |
|
26 |
Perschel |
h=1.3+b11+(b2DBHb3)−1 |
[44] |
|
27 |
Sibbesen |
h=1.3+b1DBHb2DBH−b3 |
[45] |
where, h is tree height (m), DBH is diameter at breast height (cm), b1, b2 and b3 are regression parameters.
Table 2. Model performance criteria
|
No. |
Function Name |
Equation |
|
1 |
Root mean square error (RMSE) |
RMSE=√∑(hi−ˆhi)2n |
|
2 |
Mean absolute percentage error (MAPE) |
MAPE=100×∑|hi−ˆhiyi|/n |
|
3 |
Coefficient of determination (R2) |
R2=1−∑(hi−ˆhi)2∑(hi−ˉh)2 |
|
4 |
Adjusted Coefficient of determination (R2 adj.) |
R2adj.=1−(1−R2)(n−1)(n−k) |
|
5 |
Akaike information criterion (AIC) |
AIC=2k+nln∑(hi−ˆhi)2n |
|
6 |
Bayesian information criterion (BIC) |
BIC =klnn+nln∑(hi−ˆhi)2n |
|
7. |
Mean Difference (MD) |
MD=∑ni=1(hi−ˆhi)n |
where, hi is the observed height for tree i, ˆhi is the predicted height for tree i, ˉh is the observed mean tree height, k is number of model parameters, and n is number of observations.
Figure 1. Scatter plot of height against diameter at breast height of A. macrophyllus for model fitting (a) and validation (b)
Traditional ordinal ranks for m techniques include 1,2,…, m. The models are shown in order, although not in their exact positions relative to each other. Following Poudel and Cao [46], the relative rank method is used as follows:
Ri=1+(m−1)(Si,j−Smin
where, R_i is the relative rank model i(1,2, \ldots, m), S_i is the goodness of fit statistics produced by the model i, S_{\min } is the minimum value of S_i, and S_{\max } is the maximum value of S_i. Using this technique, we only apply for models with the same character, the RMSE, MAPE, AIC, and BIC, that the smaller the value the better. Then we sum the R_i to find the best model. The ranks of each model in the five performance categories were then totalled. The model with the lowest sum was deemed the best nonlinear growth function for fitting the data.
Poudel and Cao [46] argued that in this ranking system, the best and worst approaches are ranked 1 and m, respectively. The ranks of the remaining methods are expressed as real numbers ranging from 1 to m. Because the new ranking approach considers the size as well as the order of the Sis, it should provide more information than traditional ordinal rankings. For example, relative ranks of 1, 1.3, 4.5, 4.7, and 5 in the instance of five methods indicate that the methods are divided into two groups; the first group (1 and 1.3) and the second group (4.5, 4.7, and 5) were separated by a significant gap.
3.1 Data characteristics
Table 3 indicates that the most numerous red jabon observed in this study have DBHs of 10 to just under 15 cm and heights of 10 to 15 meters. The smallest number of trees had DBHs and heights of 20 and 20 m, respectively. The correlation between diameter and tree height is very strong in both data for model development and validation (Figure 1).
Table 3. Descriptive statistics of model fitting and validation data set
|
Height (m) |
||||||||||||
|
DBH (cm) |
Model Fitting Data Set |
Validation Data Set |
||||||||||
|
<5 |
5-<10 |
10-<15 |
15-<20 |
20- |
Total |
<5 |
5-<10 |
10-<15 |
15-<20 |
20- |
Total |
|
|
<5 |
277 |
17 |
|
|
|
294 |
71 |
2 |
|
|
|
73 |
|
5-<10 |
259 |
431 |
23 |
|
|
713 |
66 |
108 |
4 |
|
|
178 |
|
10-<15 |
|
311 |
493 |
13 |
|
817 |
|
81 |
123 |
|
|
204 |
|
15-<20 |
|
|
180 |
232 |
|
412 |
|
|
40 |
64 |
|
104 |
|
20- |
|
|
5 |
124 |
4 |
133 |
|
|
4 |
28 |
1 |
33 |
|
Total |
536 |
759 |
701 |
369 |
4 |
2369 |
137 |
191 |
171 |
92 |
1 |
592 |
Table 4. Estimated parameters and statistics fits for each h-d model of A.macrophyllus in Indonesia
|
Model |
b1 |
b2 |
b3 |
MAPE |
RMSE |
R2 |
R2-adj. |
AIC |
BIC |
Rank |
|
1 |
0.626 |
1.067 |
|
14.34 |
1.26 |
0.918 |
0.918 |
1084 |
1095 |
10.2 |
|
2 |
-2.139 |
-0.151 |
|
11.13 |
1.12 |
0.934 |
0.934 |
551 |
562 |
5.1 |
|
3 |
-436.172 |
-597.265 |
|
15.52 |
1.27 |
0.915 |
0.915 |
1152 |
1164 |
11.2 |
|
4 |
28.575 |
13.696 |
|
10.35 |
1.13 |
0.933 |
0.933 |
582 |
594 |
5.1 |
|
5 |
28.575 |
-13.696 |
|
10.35 |
1.13 |
0.933 |
0.933 |
582 |
594 |
5.1 |
|
6 |
0.62 |
-0.069 |
|
14.48 |
1.26 |
0.917 |
0.917 |
1093 |
1104 |
10.3 |
|
7 |
1314.457 |
0.001 |
|
75.60 |
6.91 |
0.915 |
0.915 |
9163 |
9174 |
108.0 |
|
8 |
3.404 |
-14.998 |
|
9.98 |
1.12 |
0.935 |
0.935 |
526 |
538 |
4.5 |
|
9 |
0.73 |
-1.002 |
|
15.56 |
1.28 |
0.915 |
0.915 |
1155 |
1167 |
11.2 |
|
10 |
0.729 |
-0.002 |
|
15.53 |
1.27 |
0.915 |
0.915 |
1153 |
1164 |
11.2 |
|
11 |
3.304 |
-12.505 |
|
10.86 |
1.15 |
0.930 |
0.930 |
656 |
668 |
5.8 |
|
12 |
27.229 |
-12.505 |
|
10.86 |
1.15 |
0.931 |
0.931 |
656 |
668 |
5.8 |
|
13 |
0.551 |
2.977 |
|
11.83 |
1.16 |
0.930 |
0.930 |
689 |
701 |
6.4 |
|
14 |
0.501 |
1.644 |
|
9.90 |
1.11 |
0.936 |
0.936 |
503 |
514 |
4.3 |
|
15 |
0.5 |
0.843 |
|
69.91 |
5.53 |
0.884 |
0.884 |
8108 |
8120 |
93.3 |
|
16 |
23.602 |
153.155 |
-1.857 |
10.04 |
1.10 |
0.937 |
0.937 |
464 |
476 |
4.1 |
|
17 |
7.317 |
0.203 |
0.039 |
9.96 |
1.10 |
0.937 |
0.937 |
468 |
479 |
4.1 |
|
18 |
16.182 |
16.294 |
0.258 |
12.00 |
1.14 |
0.933 |
0.933 |
607 |
619 |
5.9 |
|
19 |
18.102 |
0.011 |
1.694 |
10.49 |
1.10 |
0.937 |
0.937 |
475 |
486 |
4.4 |
|
20 |
20.452 |
0.099 |
2.152 |
9.98 |
1.10 |
0.937 |
0.937 |
462 |
473 |
4.1 |
|
21 |
18.134 |
3.969 |
0.149 |
10.53 |
1.11 |
0.936 |
0.936 |
480 |
491 |
4.4 |
|
22 |
-385.385 |
384.926 |
0.003 |
28.87 |
3.01 |
0.918 |
0.918 |
5228 |
5240 |
48.6 |
|
23 |
3.525 |
-18.351 |
2.245 |
10.00 |
1.11 |
0.936 |
0.936 |
479 |
491 |
4.2 |
|
24 |
48.855 |
8.195 |
-0.644 |
9.99 |
1.11 |
0.936 |
0.936 |
497 |
508 |
4.3 |
|
25 |
-2.551 |
1.814 |
-0.025 |
10.24 |
1.11 |
0.936 |
0.936 |
478 |
489 |
4.3 |
|
26 |
23.602 |
0.007 |
1.857 |
11.32 |
1.15 |
0.937 |
0.937 |
676 |
688 |
6.1 |
|
27 |
0.029 |
3.96 |
-0.215 |
9.81 |
1.11 |
0.937 |
0.937 |
501 |
512 |
4.3 |
where, b1, b2 and b3 are regression parameters, MAPE is mean absolute percentage error (%), RMSE is root mean square error (m), R2 is coefficient of determinastion, AIC is akaike information criterion, BIC is Bayesian information criterion, MD is Mean Difference.
3.2 Model development and validation
At the modelling stage, the models developed have very high accuracy. This is shown by the coefficient of determination of more than 88% for all models used. The models have a coefficient of determination between 0.887 to 0.937, either with two parameters or three parameters. Based on this fact, it shows that the diameter variable can explain the diversity of tree height by more than 88% and the rest is explained by variables other than diameter (Table 4).
Based on this MAPE value, most models are categorized as good forecasting with a range of 10-20% [47], while there are 3 models that have quite different MAPE values above 20%, namely Models 7, 15, and 22. On the other hand, several models show values below 10, which indicates highly accurate forecasting [47], namely Models 27, 14, 17, 8, 20, and 24, respectively, from the smallest value.
Model 16 demonstrated a high level of prediction accuracy with a MAPE of 10.04%. The RMSE of 1.10 further confirms the precision of the model in estimating the values. Both the R² and adjusted R² were 0.937, indicating that the model accounted for 93.7% of the variability in the outcome variable. Additionally, the AIC and BIC values were 464 and 476, respectively, the lowest among all the models, suggesting an optimal balance between data fit and model simplicity. With a rank of 4.1, Model 16 was among the top models in general.
Model 17 achieved a MAPE of 9.96%, which was marginally superior to the performance of Model 16. The RMSE was 1.10, which is the same as that of Model 16. The values of R² and adjusted R² for Model 16 are both 0.937. The AIC and BIC values for this model are slightly higher than those of Model 16, with values of 468 and 479, respectively. However, they remain highly competitive. The rank of 4.1 suggests that it is equal to Model 16 in terms of performance.
Model 20 has a MAPE of 9.98%, which is very identical to the MAPE of Model 17. The RMSE value is 1.10, which is identical to the RMSE values of Models 16 and 17. The values of R² and adjusted R² for Models 16 and 17 are both 0.937. The AIC and BIC values are 462 and 473, respectively. These values represent the lowest AIC and BIC, reflecting the optimal balance of the model. The rank of this model is 4.1, which places it among the top performers alongside Models 16 and 17.
Model 23 has a MAPE of 10.00%, which is still considered good. The RMSE of this model was 1.11, which was slightly greater than that of the top three models. The values of R² and adjusted R² are both 0.936, which is slightly lower than the top three models. The AIC and BIC values are 479 and 491, respectively, which are higher than the values of the top three models. The rank of this model is 4.2, which suggests a somewhat inferior overall performance when compared to Models 16, 17, and 20.
Model 27 and 14 have a MAPE of 9.81% and 9.90%, respectively, which are the lowest among all models. This indicates that Model 14 has the highest level of prediction accuracy. The RMSE of Model 14 is 1.11, which is comparable to Model 23 but more than Models 16, 17, and 20. The values of R² and adjusted R² for Model 23 are both 0.936. The AIC and BIC values for the top models are 503 and 514, respectively, which are the highest. This suggests that there is a trade-off between the goodness of fit and the complexity of the models. With a rank of 4.3, it demonstrates strong performance but falls significantly behind the others in the total ranking.
Figure 2. Mean prediction errors in the different DBH classes (5 cm intervals) and overall MD for validation of nonlinear models of A. macrophyllus in Indonesia
The remaining 20% of the dataset was used to validate the 27 models. The mean diameter (MD) was calculated for each DBH class, with 5 cm intervals, as well as the entire dataset (Figure 2). For small-diameter classes (< 20 cm), the MD was relatively low, ranging from -1 to 0.5 meters. However, for the larger diameter class (20- cm), MD increases, with the highest underestimation being -13.29 meters in M7. Overall, the MD ranges from -6.4 to 0.05. Specifically, the value of MD for M20, M16, and M17 are -0.03, -0.05, and -0.03, respectively.
The calculation of relative rank values indicates that the best equations are found in Models 16, 17, and 20. All of these models are three-parameter models. These models exhibit consistent performance across all ranges with little overall errors. The M17 and M20 models are dependable and well-balanced, with a modest advantage over the M16 in terms of overall performance. Model 23 exhibits strong performance overall, although it does have a relatively higher negative error in the DBH range of 20-, which significantly diminishes its desirability when compared to the top three models. Model 14 exhibits commendable overall performance, although it displays some degree of unpredictability within some ranges, notably in intervals below 5 and between 10 and less than 15. Based on the new data, Models 17 and 20 demonstrate high consistency and minimum faults across all ranges, making them the top performers overall.
Three-parameter models' improved flexibility and capacity to more precisely reflect the complexity of tree development dynamics help to explain their better performance than two-parameter models. Three-parameter models introduce a third parameter that lets the nonlinear link between height and diameter at breast height (DBH) be more subtly represented. Since more exact projections of tree growth are necessary for planning thinning operations, estimating timber outputs, and optimizing replanting techniques, this enhanced precision is absolutely vital for silviculture activities and forest management. Using three-parameter models will help forest managers make more informed decisions about tree selection and breeding, therefore improving the utilization of resources and the preservation of forest ecosystems. Moreover, these models can provide early identification of any problems and growth trends, thereby enabling quick interventions to improve the condition and output of the forest stands.
Figure 3 presents the projected height as determined by the DBH using the three most accurate models developed. The results showed that all models provided similar predictions for trees with a DBH of 20 cm or less. For trees with a DBH of more than 20 cm, the models exhibited distinct patterns of height growth. This variation may be due to the limited number of observations available for larger trees.
The results of the study imply that the developed growth models could be used to assess the projected height growth of the red jabon plantation using the diameter growth, which in turn is attributed to estimating stand volume, biomass, and carbon. In the field of tree breeding, the developed growth model could be used as a basis to predict the age-age genetic correlation of red jabon in BSO in determining an effective age for genetic selection. The earlier genetic selection is applied, a higher increase of accumulative genetic gain would be obtained. In addition, the growth model indicated a high correlation between DBH and height, which is important to the weighting of traits for genetic selection under multiple traits index selection. However, it should be noted here that the use of different provenances as genetic resources of red jabon should be paid attention to due to a variety of growth patterns among the provenances.
The best-performing models, Chapman-Richards and Huang, have some really useful practical applications in actual forestry operations. By offering consistent estimates of tree height based on DBH measurements, easier and less expensive to acquire, these models can be used to improve the accuracy of forest inventories. Additionally, the close relationship between height and diameter allows for using only the DBH variable in estimating tree volume. These models also justifys the use of DBH as a single variable in developing red jabon allometrics for biomass estimation. Using height and diameter variables simultaneously is not recommended, as it can cause autocorrelation between predictor variables. By means of silviculture techniques, the exact growth projections provided by these models can guide appropriate thinning schedules, therefore preserving tree density at levels that maximize timber output and support healthy development. By spotting superior genotypes with desired growth traits, these models can also enhance breeding initiatives, therefore hastening the creation of better tree varieties. The models might direct the choice of planting sites and species composition for reforestation and afforestation efforts, therefore enhancing the success rates and sustainability of these programs. Adoption of Chapman-Richards and Huang models in forestry operations can result in more efficient management, improved resource allocation, and increased long-term planning for forest conservation and use generally.
Figure 3. Predicted height using models 16, 17, and 20 for A. macrophyllus in Indonesia
The development and study of twenty-seven nonlinear height-diameter models of Anthocephalus macrophyllus demonstrates that most concave and sigmoidal functions may accurately reflect tree height-diameter relationships.The development and study of twenty-seven nonlinear height-diameter models of Anthocephalus macrophyllus demonstrates that most concave and sigmoidal functions may accurately reflect tree height-diameter relationships. The highest-performing models, namely Models 16 (Huang), 17 (Huang), and 20 (Chapman Richards), exhibit a high level of competitiveness since their performance is nearly indistinguishable. Models 23 and 14 closely trail behind in terms of performance. The selection among these models may ultimately depend on individual preferences for MAPE against AIC/BIC or other factors, such as the interpretability of the coefficients. For example, Model 14 has the highest MAPE, which makes it appealing for applications that require precise % accuracy. Models 16, 17, and 20 demonstrate a more favorable balance between fit and complexity, as evidenced by their AIC and BIC values. Model 14 is the most accurate in terms of prediction accuracy (MAPE). If the primary considerations are overall balance and model simplicity, Models 16, 17, and 20 are the preferred choices based on AIC/BIC. Model 20 is notable for having the lowest AIC and BIC, making it the most balanced option among these measures. According to validation data, Models 17 (Huang) and 20 (Chapman Richards) perform well overall due to consistency and low mistakes across ranges.
These results have important ramifications for methods of silviculture and forest management. Using the three-parameter models will help forest managers maximize thinning schedules, increase replanting techniques' accuracy, and therefore, improve their thinning policies. These models can also guide resource allocation and policy decisions on the sustainable development of forests. Future studies should concentrate on validating the general applicability of these models by means of their application to several species and ecological settings. Furthermore improving their practical relevance in real-world forestry operations is looking at their integration with advanced data analytics and remote sensing technology. All things considered, the ongoing improvement and use of height-diameter models have considerable potential to advance efforts at sustainable forest management and conservation.
We are grateful to The Centre for Forestry Instrument Standard Assessment, The Ministry of Environment and Forestry, and The Environmental Services of Wonogiri Regency, Central Java Province for facilitating the plots establishment and data collection.
[1] Halawane, J.E., Hidayah, H.N., Kinho, J. (2015). Prospek pengembangan jabon merah, Anthocephalus macrophyllus (roxb.) havil: Solusi kebutuhan kayu masa depan. Balai Penelitian Kehutanan Manado, Badan Penelitian Pengembangan dan Inovasi, Kementerian Lingkungan Hidup dan Kehutanan.
[2] Singh, M., Kumar, P., Singh, H., Kumar, A.K.A., Kumar, R., Kumar, R. (2023). Neolamarckia cadamba: A comprehensive review on its physiological, ecological, phytochemical and pharmacological perspectives. Ecology, Environment and Conservation, 29: 241-250. http://doi.org/10.53550/EEC.2023.v29i02s.042
[3] Krisnawati, H., Kallio, M., Kanninen, M. (2011). Anthocephalus Cadamba Miq.: Ecology, Silviculture and Productivity. CIFOR.
[4] Irawan, U.S., Purwanto, E. (2014). White jabon (Anthocephalus cadamba) and red jabon (Anthocephalus macrophyllus) for community land rehabilitation: Improving local propagation efforts. Agricultural Science, 2(3): 36-45.
[5] Martawijaya, A., Kartasujana, I., Mandang, Y.I., Prawira, S.A., Kadir, K. (1989). Atlas Kayu Indonesia, Jilid II (Indonesian Wood Atlas, Volume II). IAWA Journal.
[6] Arisandi, R., Valahatul Ihda, F., Nirsatmanto, A., et al. (2024). Characterization of lipophilic extractives in the wood of anthocephalus cadamba and anthocepalus macrophyllus from progeny trial. Journal of Wood Chemistry and Technology, 44(3): 182-191. http://doi.org/10.1080/02773813.2024.2334689
[7] Wistara, N.J., Carolina, A., Pulungan, W.S., Emil, N., Lee, S.H., Kim, N.H. (2015). Effect of tree age and active alkali on kraft pulping of white jabon. Journal of the Korean Wood Science and Technology, 43(5): 566-577. https://doi.org/10.5658/WOOD.2015.43.5.566
[8] Nair, K.S.S. (2000). Insect pests and diseases of major plantation species. Colección general Biblioteca.
[9] Cahyono, T.D., Wahyudi, I., Priadi, T., Febrianto, F., Darmawan, W., Bahtiar, E.T., Ohorella, S., Novriyanti, E. (2015). The quality of 8 and 10 years old samama wood (Anthocephalus macrophyllus). Journal of the Indian Academy of Wood Science, 12: 22-28. https://doi.org/10.1007/s13196-015-0140-8
[10] Ilyas, Y., Rombang, J.A., Lasut, M.T., Pangemanan, E.F. (2015) Pengaruh media tanam terhadap pertumbuhan bibit Jabon merah (Anthocephalus macrophyllus (Roxb) Havil) (Effect of planting media on red jabon (Anthocephalus macrophyllus (Roxb) Havil) seed growth). Cocos, 6(12). https://doi.org/10.35791/cocos.v6i12.8511
[11] Masilewi, J., Nurdin, A.S., Marasabessy, M.H., Irmayanti, L., Ashari, R. (2022). Pertumbuhan bibit jabon merah (Anthocepalus macrophyllus) pada komposisi media tanam yang berbeda. ULIN: Jurnal Hutan Tropis, 6(1): 98-104. https://doi.org/10.32522/ujht.v6i1.6729
[12] Supriyanto, S., Muslimin, M., Umar, H. (2014). Pengaruh berbagai dosis pupuk organik cair urin sapi terhadap pertumbuhan semai jabon merah (Anthocephalus macrophyllus (Roxb.) Havil) (The effect of various doses of cow urine liquid organic fertilizer on the growth of red Jabon seedlings), Jurnal Warta Rimba, 2(2).
[13] Jafar, S.H., Thomas, A., Kalangi, J.I., Lasut, M.T. (2013). Pengaruh frekuensi pemberian air terhadap pertumbuhan bibit jabon merah (Anthocephalus macrophyllus (Roxb.) Havil). (Frequency Effect on growth of seedlings water provision jabon Red (Anthocephalus Macrophyllus (Roxb.) Havil)), Cocos, 2(2). https://doi.org/10.35791/cocos.v2i2.1469.
[14] A’ida, N., Larekeng, S.H., Arsyad, M.A., Putra, R.P., Restu, M. (2019). Progeny test on plant growth of 2-year-old Jabon Merah (Anthocephalus macrophyllus Roxb. Havil.) in Gowa, South Sulawesi, Indonesia: Preeliminary Study. In IOP Conference Series: Earth and Environmental Science, 270: 012020, https://doi.org/10.1088/1755-1315/270/1/012020.
[15] Sandalayuk, D., Lahjie, A.M., Simarangkir, B., Ruslim, Y. (2020). Carbon absorbtion of Anthocephalus macrophyllus and Swietenia macrophylla. King in Gorontalo, Indonesia. Journal of Biodiversity and Environmental Science, 16(5): 24-30.
[16] Mukhdlor, A., Haqiqi, M.T., Tirkaamiana, M.T., Suwinarti, W., Amirta, R. (2021) Assessment of wood biomass productivity from Anthocephalus macrophyllus forest plantation for energy production. In Proceedings of the Joint Symposium on Tropical Studies (JSTS-19), Indonesia. https://doi.org/10.2991/absr.k.210408.005
[17] Larekeng, S.H., Restu, M., Gusmiaty, G., Millang, S., Bachtiar, B. (2018). Moderate level of genetic diversity in Anthocephalus macrophyllus Roxb, an endemic tree of Sulawesi and its implication in conservation. International Journal of Agriculture System, 6(1): 74-81. https://doi.org/10.20956/ijas.v6i1.1449
[18] Faida, L.R.W., Marhaento, H. (2019). Extinction risk analysis of Anthocephalus macrophyllus in Gunungsewu Karst Area, Southern Java, Indonesia. Biodiversitas Journal of Biological Diversity, 20(7): 1897-1903. https://doi.org/10.13057/biodiv/d200715
[19] Lebedev, A., Kuzmichev, V. (2020). Verification of two- and three-parameter simple height-diameter models for birch in the European part of Russia. Journal of Forest Science, 66(9): 375-382. https://doi.org/10.17221/76/2020-JFS
[20] Van Laar, A., Akça, A. (2007). Forest Mensuration (Vol. 13). Springer Science & Business Media.
[21] Mensah, S., Pienaar, O.L., Kunneke, A., du Toit, B., Seydack, A., Uhl, E., Pretzsch, H., Seifert, T. (2018). Height–Diameter allometry in South Africa’s indigenous high forests: Assessing generic models performance and function forms. Forest Ecology and Management, 410: 1-11. https://doi.org/10.1016/j.foreco.2017.12.030
[22] Sharma, M., Parton, J. (2007). Height–diameter equations for boreal tree species in Ontario using a mixed-effects modeling approach, Forest Ecology and Management, 249(3): 187-198. https://doi.org/10.1016/j.foreco.2007.05.006
[23] Ng'andwe, P., Chungu, D., Yambayamba, A.M., Chilambwe, A. (2019). Modeling the height-diameter relationship of planted Pinus kesiya in Zambia. Forest Ecology and Management, 447: 1-11. https://doi.org/10.1016/j.foreco.2019.05.051
[24] Mehtätalo, L., de-Miguel, S., Gregoire, T G. (2015). Modeling height-diameter curves for prediction. Canadian Journal of Forest Research, 45(7): 826-837. https://doi.org/10.1139/cjfr-2015-0054.
[25] Surip, S., Indrioko, S., Nirsatmanto, A., Setyaji, T. (2017). Pengaruh seleksi terhadap perolehan genetik pada uji keturunan generasi pertama (F-1) jabon merah (Anthocephalus macrophyllus (Roxb.) Havil) di Wonogiri. Jurnal Pemuliaan Tanaman Hutan, 11(1): 33-44. https://doi.org/10.20886/jpth.2017.11.1.33-44
[26] Peng, C., Zhang, L., Liu, J. (2001). Developing and validating nonlinear height–diameter models for major tree species of Ontario’s Boreal Forests. Northern Journal of Applied Forestry, 18(3): 87-94. https://doi.org/10.1093/njaf/18.3.87
[27] Lappi, J. (1997). A longitudinal analysis of height/diameter curves. Forest Science, 43(4): 555-570. https://doi.org/10.1093/forestscience/43.4.555
[28] Lumbres, R.I.C., Lee, Y.J., Seo, Y.O., Kim, S.H., Choi, J.K., Lee, W.K. (2011). Development and validation of nonlinear height–DBH models for major coniferous tree species in Korea. Forest Science and Technology, 7(3): 117-125. https://doi.org/10.1080/21580103.2011.594610
[29] Aertsen, W., Kint, V., van Orshoven, J., Özkan, K., Muys, B. (2010). Comparison and ranking of different modelling techniques for prediction of site index in Mediterranean mountain forests. Ecological Modelling, 221(8): 1119-1130. https://doi.org/10.1016/j.ecolmodel.2010.01.007
[30] Huxley, J.S., Teissier, G. (1936). Terminology of relative growth. Nature, 137(3471): 780-781. https://doi.org/10.1038/137780b0
[31] Huang, S., Price, D., Titus, S.J. (2000). Development of ecoregion-based height–diameter models for white spruce in boreal forests. Forest Ecology and Management, 129(1-3): 125-141. https://doi.org/10.1016/S0378-1127(99)00151-6
[32] Curtis, R.O. (1967). Height-diameter and height-diameter-age equations for second-growth Douglas-fir. Forest Science, 13(4): 365-375. https://doi.org/10.1093/forestscience/13.4.365
[33] Meyer, H.A. (1940). A mathematical expression for height curves. Journal of Forestry, 38(5): 415-420. https://doi.org/10.1093/jof/38.5.415
[34] Wykoff, W. (1982). User's Guide to the Stand Prognosis Model. US Department of Agriculture, Forest Service, Intermountain Forest and Range Experiment Station.
[35] Bates, D.M., Watts, D.G. (1980). Relative curvature measures of nonlinearity. Journal of the Royal Statistical Society: Series B (Methodological), 42(1): 1-16. https://doi.org/10.1111/j.2517-6161.1980.tb01094.x
[36] Staudhammer, C., LeMay, V. (2000). Height prediction equations using diameter and stand density measures. The Forestry Chronicle, 76(2): 303-309. https://doi.org/10.5558/tfc76303-2
[37] Buford, M.A. (1986). Height-diameter relationships at age 15 in loblolly pine seed sources. Forest Science, 32(3): 812-818. https://doi.org/10.1093/forestscience/32.3.812
[38] El Mamoun, H.O., El Zein, A.I., El Mugira, M.I. (2013). Modelling height-diameter relationships of selected economically important natural forests species. Journal of Forest Products & Industries, 2: 34-42.
[39] Ogana, F.N. (2018). Comparison of a modified log-logistic distribution with established models for tree height prediction. Journal of Research in Forestry, Wildlife and Environment, 10(2): 49-55.
[40] Yang, R.C., Kozak, A., Smith, J.H.G. (1978). The potential of Weibull-type functions as flexible growth curves. Canadian Journal of Forest Research, 8(4): 424-431. http://doi.org/10.1139/x78-062.
[41] Huang, S., Titus, S.J., Wiens, D.P. (1992). Comparison of nonlinear height–diameter functions for major Alberta tree species. Canadian Journal of Forest Research, 22(9): 1297-1304. https://doi.org/10.1139/x92-172
[42] Larsen, D.R., Hann, D.W. (1987). Height-diameter equations for seventeen tree species in southwest Oregon. Oregon State University.
[43] Stage, A.R. (1963). A mathematical approach to polymorphic site index curves for grand fir. Forest Science, 9(2): 167-180. http://doi.org/10.1093/forestscience/9.2.167
[44] Peschel, W. (1938). Mathematical methods for growth studies of trees and forest stands and the results of their application. Tharandter Forstliches Jahrburch, 89: 169-247.
[45] Sibbesen, E. (1981). Some new equations to describe phosphate sorption by soils. Journal of Soil Science, 32(1): 67-74. https://doi.org/10.1111/j.1365-2389.1981.tb01686.x
[46] Poudel, K.P., Cao, Q.V. (2013). Evaluation of methods to predict Weibull parameters for characterizing diameter distributions. Forest Science, 59(2): 243-252. https://doi.org/10.5849/forsci.12-001
[47] Lewis, C. D. (1982). Industrial and Business Forecasting Methods: A Practical Guide to Exponential Smoothing and Curve Fitting. Butterworth Scientific, London, Boston.