Dauren Serikbay![]() | Lyudmila Zotova*
| Lyudmila Zotova*![]() | Aibek Zhumalin
| Aibek Zhumalin![]() | Aissarat Gajimuradova
| Aissarat Gajimuradova![]() | Aiman Rysbekova
| Aiman Rysbekova![]() | Firuz Abdulloyev
| Firuz Abdulloyev![]() | Liang Chen
| Liang Chen![]() | Timur Savin
| Timur Savin![]() | Tat`yana Sereda
| Tat`yana Sereda![]() | Zhangchen Zhao
| Zhangchen Zhao![]()
© 2024 The authors. This article is published by IIETA and is licensed under the CC BY 4.0 license (http://creativecommons.org/licenses/by/4.0/).
OPEN ACCESS
This study aims to assess the effectiveness of dwarfing genes Rht-B1a/B1b and Rht-D1a/D1b on the phenotype and yield of spring soft wheat under drought conditions in Kazakhstan. From 2018 to 2022, laboratory experiments were conducted at S. Seifullin Kazakh Agrotechnical Research University, and field trials were performed at two sites in Kazakhstan. The study involved phenotypic analysis of plant height (PH) and coleoptile length (CL), as well as yield measurement. The RhtB1a and RhtD1a alleles significantly influenced plant traits. Hybrids carrying the RhtB1a allele exhibited a reduction in plant height by 15-20%, while yield improvements reached up to 30% compared to non-dwarf varieties. Specific hybrids, such as WH190, demonstrated yields of 2.5 tons per hectare, which is 80% higher than the country's average yield indicator. Dwarfing genes RhtB1a and RhtD1a are critical for optimizing wheat growth and yield under drought conditions. This study highlights the potential of specific genotypes in wheat breeding programs, emphasizing the importance of integrating genetic and field data to identify optimal genotypes for specific environments.
spring wheat, drought conditions, semi-dwarf genes, coleoptile length, plant height, selection
Wheat is the most common and widely demanded crop due to its economic and social significance [1]. Cultivated on over 220 million hectares of land worldwide, wheat production is a crucial global agricultural activity. Asia contributes the largest share to global wheat production (44%), followed by Europe (34%) and the Americas (15%), with smaller but comparable proportions from Oceania and Africa (3.4-3.5%).
Kazakhstan, being one of the major suppliers of grains and grain products, plays a pivotal role in connecting the larger Central Asian region. The annual wheat production in Kazakhstan ranges from 12 to 12.5 million tons, of which only 6 million tons are utilized for domestic consumption, while the remaining portion is exported [2]. According to the Statistics Bureau of the Republic of Kazakhstan, the average crop yield per hectare in the country was 11.8 tons in 2020, 9.3 tons in 2021, 12.8 tons in 2022, and 12.3 tons in 2023 [3].
In recent years, global warming and climate change have significantly impacted the productivity of agricultural crops, including wheat. According to the 2021 report, wheat reserves in the country decreased by four million tons owing to drought [4].
FAO forecasts a decrease in the yield of spring wheat in Kazakhstan by 2030, ranging from 13% to 37%. Simultaneously, the cultivated areas are expected to decrease by 23% to 81%, with direct economic losses in the industry estimated at 456.93 billion tenge. By 2050, wheat yield losses are projected to be between 20% and 49%, leading to a reduction in the industry's gross output to 608.19 billion tenge at 2019 prices. Considering Kazakhstan's position as the 9th largest wheat producer and the 7th largest exporter globally, and the exclusive exporter in Central Asia, the lack of climate adaptation measures in Kazakhstan could pose a threat to the entire region's food security [5].
One potential tool to enhance wheat productivity in Kazakhstan is the introduction of genes associated with the "Green Revolution" into breeding programs. The Green Revolution resulted in the improvement of the wheat yield index due to a significant height reduction of the straw [6]. This was facilitated by identifying the main dwarfism genes currently widespread in world breeding programs. The decrease in height resulting from the shortening of straw by reduced height genes (Rht) has long been proposed as a factor in increasing the wheat yield index [7, 8].
Agronomic outcomes observed in lines carrying Rht-B1b or Rht-D1b alleles include a 15-20% reduction in PH and increased grain yield [7]. The presence of either Rht-B1b or Rht-D1b alone decreases PH by 14.6%, but the combined presence of Rht-B1b and Rht-D1b reduces PH by 41%. Moreover, lines containing Rht-B1b and Rht-D1b have shorter leaves compared to genotypes containing Rht-B1a and Rht-D1a, which may affect photosynthesis activity [9]. Several comparative studies have shown that wheat yield decreases with PH reduction, limiting the range of PH for optimal yield [7]. However, recent research has demonstrated that Rht-B1b and Rht-D1b significantly improve potential wheat yield when nitrogen fertilizers are used but reduce coleoptile length (CL), seedling strength, and grain weight (GW). Therefore, considerations of grain square (GS), TGW, CL, and quality indicators are necessary when selecting hybrids and varieties directly affecting both yield quantity and quality [10, 11]. In arid and semi-arid regions, coleoptile length is a crucial agronomic trait in wheat selection. The length of the coleoptile determines the maximum seeding depth, playing a decisive role in crop yield formation. Wheat varieties with longer coleoptiles can be sown at greater depths, allowing seeds to absorb sufficient moisture for germination and establishment, thereby ensuring a normal crop yield [12]. Therefore, the analysis of the genetic basis of coleoptile length is essential for improving wheat in arid and semi-arid regions. For instance, genes such as Rht1 and Rht2, as reported by Rebetzke et al. [13], reduce both coleoptile length and plant height. In the conducted QTL analysis by Xu et al. [14], it was demonstrated that the identified QTLs, QCL.qau-4BS and QCL.qau-4DS, could account for 9.1–22.2% of phenotypic variations in different environments at the Rht-B1 and Rht-D1 loci, respectively. Following transcriptome studies, the researchers identified several key pathways involving the function of Rht-B1b in coleoptile development, including phytohormones, circadian rhythm, and starch and sucrose metabolism [14]. In other studies, it has been reported that Rht-B1b may reduce wheat coleoptile length (CL), but molecular-level investigations were lacking. To confirm that the Rht-B1 gene is a functional locus for the 4B QTL, overexpression lines Rht-B1b-OE were studied, demonstrating that overexpression of Rht-B1b can decrease CL, while loss of function in Rht-B1b leads to an increase in CL compared to null transgenic plants [15, 16].
According to available data, various single nucleotide mutations in both short-stemmed genes can play a remarkable role in wheat breeding for increased productivity.
The aim of this study is to assess the effectiveness of the influence of dwarfing genes Rht-B1a/B1b, RhtD1a/D1b, on the phenotype (PH, CL, JG) and overall yield (TY) of spring soft wheat varieties and hybrids under drought conditions.
2.1 Research design
The research was conducted from 2018 to 2022, encompassing both laboratory and field experiments. Laboratory experiments were carried out at the Agricultural Biotechnology Research Platform of the S. Seifullin Kazakh Agrotechnical Research University, Astana, Kazakhstan. Field experiments were conducted at the A.F. Khristenko Karaganda Agricultural Experimental Station (Central Kazakhstan) and the "Niva" farm (Northern Kazakhstan). These areas were selected due to their distinct soil and climatic conditions, which allowed for a comprehensive assessment of the wheat varieties under study.
2.2 Parental and hybrid information
A total of 124 parental forms were selected in previous studies under soil drought conditions in both natural and artificially induced drought conditions (slow-developing drought stress and rapid dehydration) [17]. Selection was performed using single nucleotide polymorphism (SNP) technology with 19 SNP markers, genotyping of SNPs, and 18 markers for the DREB2-gene encoding transcription factors TaDREB5 and TaNFYC-A7 [18].
Hybridization was carried out using the "twel" method of limited free pollination by Borlaug [19] with the most drought-resistant parental forms (Xn03, Xn04, Xn08, Xn09, Xn10, Xn13 - China, Gladius, RAC 875, Wyalkatchem, Kite – Australia, MMF-044 – Africa, Saratovskaya 66, Lyutestcens 141 – Russia, Erythrospermum 35, Tauelsizdyk 20, Astana, Akmolinskaya 2, Aktiubinka, Karabalykskaya 25, 90, Karagandinskaya 22, 25, 29, 30, 31, Shortandinskaya, 2012, 2017 - Kazakhstan) based on phenotypic indicators under drought conditions (2018). After the study of 167 hybrids 20 promising ones were selected for further investigation. The hybrid nursery was established following the scheme of maternal, hybrid, and paternal forms, with each sample's plot area being 1 m². The research collection was sown annually in two agroecological points, and samples were collected from a 2 m² area. Additional information on parental forms is presented in Table S1.
Control setups included non-dwarf wheat varieties and parental lines without the Rht genes under study. These controls were crucial for comparing the phenotypic traits and yield performance of the hybrids.
2.3 Field experiment conditions
All field experiments were conducted in two agroecological points in Central and Northern Kazakhstan. Soils in both regions are dark chestnut. The climate is characterized by a sharply continental climate. The climate was arid, and in the last 5 years, climatic conditions under conditions of global warming led to an increase in the average annual temperature and a decrease in total precipitation [20]. According to the hydrothermal index by Selyaninov [21], in the central part of the country, it was 0.72 in 2018, 0.67 in 2019, 0.63 in 2020, 0.33 in 2021, and 0.39 in 2022. A similar indicator in Northern Kazakhstan in 2018 was 0.72, in 2019 - 0.42, in 2020 - 0.89, and in 2021 and 2022 - 0.34 and 0.39, respectively. During the experiments, climatic conditions in both locations were dry and very dry.
2.4 Weather conditions
Monthly weather conditions in Central and Northern Kazakhstan are reported by the National Hydrometeorological Service of Kazakhstan - Kazhydromet [22]. Analyzed and averaged temperature and precipitation data from 2018 to 2022 are presented in Figure 1.
2.5 Phenotyping parameters of parental forms and hybrids
2.5.1 Measurement of PH
To determine PH, 20 wheat plants were randomly selected from the center of each plot. Using a ruler, the height of each plant from the ground to the tip of the spike was measured (accuracy 0.5 mm), and the average height of 20 plants was taken as PH [23].
A) Temperature indicators in Central Kazakhstan and in Northern Kazakhstan
B) Precipitation indicators in Central Kazakhstan and in Northern Kazakhstan
Figure 1. Weather conditions in the period 2018–2022 in the Central Kazakhstan regions and Northern Kazakhstan, A) – temperature parameters, B) – precipitation level
Table 1. Types of primers used on experiments
|
Name of Gene |
Nucleotide Sequence of Primers |
Product Size |
Reference |
|
Rht- B1b-F |
5′-GGTAGGGAGGCGAGAGGCGAG-3′ |
237 bp |
[8] |
|
Rht- B1b-R |
5′-CATCCCCATGGCCATCTCGAGCTA-3′ |
||
|
Rht- B1а-F |
5′-GGTAGGGAGGCGAGAGGCGAG-3′ |
237 bp |
|
|
Rht- B1а-R |
5′-CATCCCCATGGCCATCTCGAGCTG-3′ |
||
|
Rht-D1b-F |
5′-CGCGCAATTATTGGCCAGAGATAG-3′ |
254 bp |
|
|
Rht-D1b-R |
5′-CCCCATGGCCATCTCGAGCTGCTA-3′ |
||
|
Rht-D1a-F |
5′-GGCAAGCAAAAGCTTCGCG-3′ |
264 bp |
|
|
Rht-D1a-R |
5′-GGCCATCTCGAGCTGCAC-3′ |
2.5.2 Measurement of CL
For CL measurement, 15 seeds of uniform size and without damage were selected. The seeds were sown in soil in plastic trays, placed in a completely dark box, and kept in a growth chamber at a constant temperature of 22℃. After 10 days, the average CL of 10 seedlings was measured in millimeters from the base of the seed to the tip of the coleoptile [24].
2.5.3 Measurement of JG index
To calculate the JG index (Galchenko) in field conditions, 10 fully ripe plants were picked, and measurements of the length of the second internode and the external diameter were taken using an electronic caliper (Matrix, 31611). After obtaining the average value of each parameter, the index was presented as a percentage [25].
2.5.4 Yield parameter
The TY was determined by collecting plants from a 1 m² plot in duplicate. The grains were separated from the spikes using the WinterSteiger LD350 thresher. Subsequently, the threshed and cleaned seeds were weighed on OHAUS Pioneer PA114C scales with an accuracy of 0.1 g. The average weight from the two plots was considered as the TY value.
2.6 Gene Identification by PCR
Plant material in field conditions was collected after reaching the flowering stage. Two leaves were collected from each plant. DNA extraction was performed using the CTAB method [26]. All genes were amplified using the VeritiPro amplifier (Applied Biosystems, USA). The primer sequences are presented in Table 1. PCR reaction mixture and conditions for each gene are presented in Tables 2-4.
Table 2. The PCR reaction mixture (total volume 20 µl)
|
Mixture Component |
Concentration |
Volume, µl |
|
1 × DreamTaq Buffer |
10х |
1.5 |
|
dNTPs |
4 nmol |
1.5 |
|
Primer F |
10 pmol |
0.5 |
|
Primer R |
10 pmol |
0.5 |
|
DreamTaq Hot Start DNA Polymerase (Thermo Fisher Scientific, USA) |
1.25 U |
0.2 |
|
DNA matrix |
100 ng |
2 |
|
H2O |
mQ |
3.8 |
Table 3. PCR conditions for the Rht-B1a Rht-B1b genes
|
Reaction Stage |
Temperature, ℃ |
Time |
Number of Cycles |
|
Initial Denaturation |
95 |
5 min |
1Х |
|
Denaturation |
95 |
30 sec |
30Х |
|
Primer Annealing |
65 |
30 sec |
|
|
Extension |
72 |
30 sec |
|
|
Final Extension |
72 |
1 min |
1Х |
Table 4. PCR conditions for the Rht-D1a Rht-D1b genes
|
Reaction Stage |
Temperature, ℃ |
Time |
Number of Cycles |
|
Initial Denaturation |
95 |
5 min |
1Х |
|
Denaturation |
95 |
20 sec |
45Х |
|
Primer Annealing |
58 |
30 sec |
|
|
Extension |
72 |
10 sec |
|
|
Final Extension |
72 |
2 min |
1Х |
PCR products were separated on a 2% agarose gel and visualized after ethidium bromide staining using the Doc-Print imaging system (Vilber Lourmat, France) [9].
2.7 Statistical analysis
The gathered data underwent scrutiny to assess normality and consistent variance. All experiments were carried out in three replications. Data analyses were conducted using the GLIMMIX procedure of SAS (Version 9.4, SAS Institute Inc., Cary, NC) using the mixed procedure with block as the random factor. The least squares mean statement in SAS with the Tukey adjustment at P=0.05 was used for comparison of means. The p-value was calculated relative to the mother and father of the hybrid. The phenotypic and genetic indicators were analyzed using the SPSS version 22.0 program with a nonparametric (Man-Whitney test). A Spearman correlation analyze was conducted to evaluate the relationships between the dwarf-genes and productivity genes.
3.1 Phenotype analyses
3.1.1 Plant height
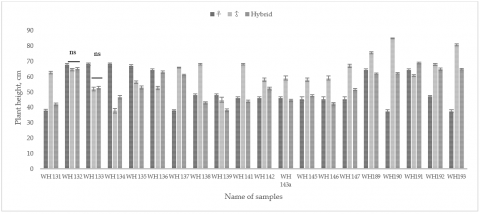
PH in field conditions in two agroecological zones was the most variable parameter. Depending on the weather conditions in the two zones, the difference in PH was approximately 10-20 cm. The hybrids demonstrated consistent plant height under drought conditions, indicating the effectiveness of dwarfing genes. Chinese varieties had the lowest PH levels (40-53 cm), decreasing by 1-13 cm in the central part of the country. The hybrids in Central Kazakhstan exhibited a lower plant height compared to the Northern region, ranging from 0.9 to 18%, with the WH147 hybrid being 6% taller. The five-year average values of PH for maternal forms were 51.6±0.98 cm (p<0.01), paternal forms - 62.3±2.5 cm (p<0.05), and hybrid forms - 53.5±2.2 cm (p>0.05) [27]. This reduction in PH is crucial for improving wheat yield under drought conditions, as shorter plants are less prone to lodging and more efficient in water use. Figure 2 shows the average differences in PH between parental and hybrid forms in field conditions.
According to the manifestation of characteristics, for 9 hybrids, PH from 37 cm to 64.2 cm (7-11%) was higher than that of the maternal form. For 11 hybrids, the PH level was lower by 2.7 to 9.8 cm (4-20%) compared to the maternal form. At the same time, paternal lines were higher for 15 hybrids by 0.5-22 cm (0.7-27%). Only 5 hybrids (25%) were identified as taller hybrids compared to paternal forms. Only the WH191 hybrid was higher than both parents by 8.1 cm of the maternal form and by 4.7 cm of the paternal form. Among the dwarf hybrids, WH131, WH133, WH134, WH138, WH139, WH141, WH142, WH143a, WH145, WH146, and WH147 stood out, with an average PH ranging from 38.26±0.3 to 53±0.4 cm. The observed characteristics of the chosen parental forms demonstrated their effectiveness. Hybridizing semi-dwarf Chinese and drought-resistant Australian varieties with tall varieties from Kazakhstani and Russian breeding resulted in hybrids demonstrating consistent plant height under drought conditions in both regions.
According to Islam et al. [28] during a three-year study of the height of cereal crops the average height of the plant was 97 cm in 2008–2009, 87 cm in 2009–2010, and 91 cm in 2010–2011. Rye had the tallest plants (height ranged from 100–110 cm), followed by triticale lines/varieties (85–105 cm) and wheat (60–88 cm). The average stem diameter for the three growing seasons was 0.31 cm in the rye, 0.32 cm in triticale, and 0.24 cm in wheat [24].
Figure 2. Difference in PH of parent forms and hybrids in the field of spring wheat (2018-2022)
nsp≥0.05, all not marked samples have p<0.001.
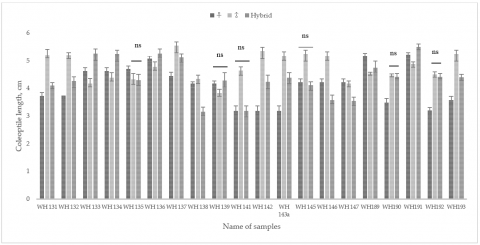
Figure 3. The average CL of parental forms and hybrids (2018-2022)
nsp≥0.05, not marked samples have p<0.001.
Gao et al. [29] found that irrigation at the stage of tube entry stage increases the height of wheat plants by 6.60–9.70%. Compared with wild-type alleles Rht-B1a and Rht-D1a, the genetic background of Rht-B1b and Rht–D1b significantly reduced plant height by 14.40–15.50% and 16.90-19.10%, respectively. Rht-B1b and Rht-D1b can reduce plant height by 29.10–32.50% [29]. Plant height is notably negatively correlated with grain yield during irrigation, whereas plant height is positively correlated with the drought coefficient.
3.1.2 Measurement of CL
Seed placement depth during drought is critical for uniform seedlings, making CL one of the pivotal parameters in the selection of hybrids in this study. A long coleoptile is a highly beneficial trait for arid zones. Figure 3 presents average CL values for parental forms and hybrids.
CL played a crucial role in the selection of hybrids in this study, particularly considering the annual rise in drought conditions. Thus, based on the CL in 60 samples the average value in maternal varieties was 4.1±0.15 cm (p<0.01); in paternal varieties, the CL was 4.8± 0.1 cm (p<0.05), and in hybrids, the CL was 4.4±0.15 cm (p>0.05) (Figure 2). In 60 samples, the average CL for maternal varieties was 4.1±0.15 cm (p<0.01); for paternal varieties, CL was 4.8±0.1 cm (p<0.05), and for hybrids, CL was 4.4±0.15 cm (p>0.05). Samples with CL exceeding 5 cm were identified among hybrids WH191, WH133, and WH134. Among maternal varieties, Xn-08 had the most significant long coleoptile; among paternal varieties - Astana, Aktyubinsk, Karagandinskaya 30, Tauelsizdyk 20, Xn-09, and Lyutescens 141. Long coleoptiles are mainly observed in paternal forms (60%), while 25% of hybrids (WH133, WH134, WH136, WH139, WH191) are taller than both parents, and 30% of hybrids (WH135, WH138, WH141, WH145, WH146, WH147) are shorter than both parents. At the same time, if CL of paternal forms is at least 5 cm or more, the length of the hybrid reaches 4 cm or more (WH131, WH132, WH142, WH143a, WH193). In 40% of samples, CL in hybrids was 4 cm or more, and the CL in both parental forms exceeded 4 cm and 5 cm, respectively. As a result, varieties from Chinese (Xn-04, 08, 09, 10, 13), Kazakhstani (Karabalikskaya 25, Tauelsizdyuk 20, Astana, Shortandinskaya 2017, Akmola 2, Aktubinka, Karagandinskaya 29 and 22, 30 and 31, Karabalikskaya 90), and Russian (Saratovskaya 66, Lyutescens 141) breeding possess optimal CL parameters for our region.
The correlation analysis showed a strong positive relationship between CL and drought resistance, with longer coleoptiles enhancing seedling emergence and establishment under water-deficient conditions.
For instance, according to measurements of CL in the arid climate of Australia, which typically ranges from 5.2 to 7.3 cm, the importance of CL exceeding 5 cm is highlighted [30]. According to Rebetzke et al. [31], varieties with short coleoptiles (6.5-10.2) had lower viability with deep planting (30-40%), whereas varieties with long coleoptiles of 13.2 cm and 15.1 cm showed viability up to 80%. For the studied regions of Kazakhstan, CL values ranging from 4 to 6 cm are considered optimal.
3.1.3 JG index
The JG index determines the resistance to lodging, which depends on the morphological parameters of the stem. A comprehensive assessment allows one to assess the available varieties and lines more accurately. For this purpose, the JG index l2 / d2 is also determined, the ratio of the length of the second internode to its diameter [32].
Samples are deemed resistant to lodging when the index value is below 35%, moderately resistant when it falls between 35-45%, and lodging-prone if it surpasses 45%. Plants with reduced stem height and increased thickness are regarded as resilient to lodging, particularly in arid regions where stem strength plays a crucial role in determining TY of crop during water-deficient conditions.
According to the results presented in Table S2, all samples, both parental and hybrids, demonstrated resistance to lodging. The average lodging resistance for the Saratovskaya 66 and Astana varieties (36.05±0.14) was the highest. The mean JG ratio for maternal varieties was 25.6±1.0 (p<0.01), for paternal varieties - 29.9±0.9% (p<0.01), and among hybrids, it was 25.69±0.9 (p>0.05). All samples from maternal forms and hybrids had a JG parameter below 35%; only in paternal forms, for the Astana variety, the JG index reached 36.05±0.14. Varieties with the lowest index were RAC 875 (19.1±0.3), Wyalkatchem (21.9±0.3), and WH133 (13.1±0.2). Based on these results, all samples were classified as resistant to lodging.
3.2 Yield parameters
The genetic makeup governing grain TY is intricate, relying on multiple interconnected traits. Furthermore, the high level of genotype interaction with the environment and the low heritability of this ultimate trait, in most cases, render the selection process highly challenging. In such a situation, indirect selection of TY based on highly heritable correlated traits [33] would be more practically feasible. Figure 4 presents the results of TY for parental forms and hybrids.
Only 15% of hybrid samples exhibited a yield of 2 tons or more per hectare, while among the maternal forms, only 40% had yields exceeding 2 t/ha, and among the paternal forms, only 25% achieved a similar yield. For the WH190 hybrid, the yield reached 2.5 t/ha, which is 80% higher than the country's average multi-year yield indicator. According to genetic analysis, maternal line samples with high yield had RhtD1b and RhtBa alleles in the genome of Chinese selection varieties Xn-02, Xn-03, Xn-10, Xn-13. In other samples, the presence of one of the genes RhtD1b, RhtB1b, RhtD1a was observed in the genome. The genetic picture for paternal lines is diverse: the Xn-09 sample has all 4 alleles in the genome, the Kazakh selection variety Akmola 2 has 3 alleles Rht B1a, Rht D1a, Rht B1b, and the Karaganda 22 variety has 2 alleles Rht B1a, Rht D1a. The Astana and Karaganda 30 varieties have one allele each of RhtD1b and Rht B1a, respectively. Hybrid samples WH137, WH190, and WH192 have yields exceeding 2 t/ha, and it is important to note that the parental forms of these hybrids all have yields of 2 tons or more per hectare. The maternal lines are varieties of Chinese selection with high yields, and the paternal form is a variety of Kazakh selection with high yields.
For the arid climate of Northern and Central Kazakhstan, semi-dwarf varieties of Chinese selection appear to be the most promising, both as independent breeding material and as gene donors for Rht genes in breeding programs. This measurement is deemed sufficiently elevated for areas characterized by a dry climate. Samples with parental forms carrying both productivity genes showed the highest results in TY.
Figure 4. Productivity of parental and hybrids forms wheat, 2018-2022
nsp≥0.05, ***p<0.001.
3.3 Genetic parameters
Samples were scrutinized at the genetic level with the aim of investigating parental forms and hybrids, specifically focusing on semi-dwarfism and coarseness traits. To accomplish this, we examined genes related to the "green revolution," specifically Rht-B1a, Rht-B1b, Rht-D1a, and Rht-D1b, which are widespread in almost all modern varieties.
3.3.1 Rht-genes
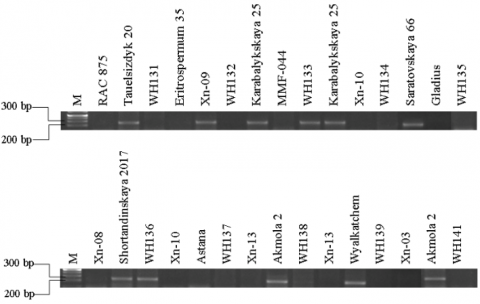
Table S1 displays the average values of all investigated parameters that determine features like short stature and lodging resistance (PH and JG), along with CL. Figure S1 demonstrates the results of PCR for the identification of Rht-genes. According to molecular-genetic analysis, dwarf varieties contain at least one of the Rht-B1b and Rht-D1b-genes in their genome. Australian varieties carried a single Rht-D1b-gene with average values of PH 46.42 cm and CL 3.96 cm. The Chinese variety Xn-09 had all four alleles of the Rht-gene, with PH at 64.74±0.87 cm and CL at 5.19±0.06 cm. Chinese samples are highly appropriate for cultivation in arid regions, encompassing both Central and Northern Kazakhstan, owing to their optimal equilibrium between PH and CL. As for Russian and Kazakh selection varieties, they contained one to two genes of Rht-B1a and Rht-D1a with average values of PH 67.17±0.3 cm and CL 4.8±0.05 cm. This ensures the presence of Rht-B1b and Rht-D1b in Russian and Kazakh samples. Nevertheless, the presence of Rht-D1b or Rht-B1b genes in the genetic makeup of Astana, Akmola 2, and Shortandinskaya 2012 varieties results in PH values ranging from 66 to 68 cm and CL values between 4.18 and 5.54 cm, which are well-suited for the climatic conditions of Kazakhstan.
Thus, hybrids WH 131, WH 132, WH 133, WH 134, WH 135, WH 137, WH 138, WH 139, WH 141, WH 142, WH 145, and WH 193 had both positive and negative alleles in their genome; the average PH was 50.9±0.3, with CL at 4.3±0.07. Among them, hybrids WH 133, WH 134, and WH 137 had CL exceeding 5 cm. Analysis of the data uncovered that the occurrence or absence of dwarfing genes Rht-B1b/Rht-D1b doesn't necessarily ensure the absence or presence of the Rht-B1a/Rht-D1a-gene, as seen in varieties Erithrospermum 35, Xn-08 and Akmola 2, for instance. However, it leads to a phenotype characterized by a short stem, attributed to the presence of the Rht-D1b-gene (as observed in Wyalkatchem and WH 147). Nevertheless, sample Xn-09 carries four genes, which may manifest differently under various climatic conditions. Hybrids WH131-135, WH137, WH139, WH141-142, WH145, WH147, WH193 inherited the Rht-D1b gene from parental forms (varieties from China, Africa, Australia). These hybrids can be considered semi-dwarf based on 5-year trials. In this case, CL for these hybrids ranged from 4.1-5.12 cm, except for WH147 with CL at 3.14 cm. The existence of a singular gene with a positive influence can lead to a reduction in length without causing a substantial negative impact on CL. Therefore, the presence of the Rht-D1b-gene is preferable for long-culm hybrids with a shortened stem.
3.4 Gene correlation
3.4.1 The Maine-Whitney correlation analysis
In accordance with the Maine-Whitney correlation analysis, there is a direct correlation between the presence of short-stem genes and the phenotypic index of CL. To evaluate the influence of different genetic allelic variations on agronomic traits, information from 33 parental materials was utilized to analyze the genetic effects of Rht-B1b, Rht-D1b (Table 5). Statistical analysis was conducted using SPSS software version 22.0, employing nonparametric tests.
Table 5. Effect of Rht-D1 and Rht-B1 on CL, PH and JG
|
|
N |
CL (cm) |
PH (cm) |
JG |
TY (t/ga) |
|
Rht-D1a |
16 |
4.81±0.44 |
66.74±8.21 |
27.83±5.49 |
1.85±0,07 |
|
Rht-D1b |
17 |
4.1±0.64*** |
48.72±9.66*** |
27.5±4.49 |
1.8±0,05 |
|
Rht-B1a |
28 |
4.44±0.66 |
56.62±13.15 |
27.57±5.01 |
1.9±0,05 |
|
Rht-B1b |
5 |
4.43±0.69 |
62.17±9.78 |
28.17±4.92 |
1.6±0,1** |
Note: *** denote significance between genotypes at P-value 0.001, ** P-value 0.01.
As evident from the results of the table, the maximum level of total yield is observed in the genotype with the alleles Rht-B1a and Rht-D1a, ranging from 1.85 to 1.9 t/ha. Additionally, these same genotypes exhibited optimal values for the parameters CL (coleoptile length) at 4.43-4.81 cm, PH (plant height) at 56.62-66.74 cm, and JG (germination index) at 27.57-27.83%. Simultaneously, Rht-D1b apparently reduced PH and shortened CL by 51.67% and 30.45% (р<0.001), respectively.
3.4.2 Spearman correlation
A Spearman correlation analysis were conducted to evaluate the relationships between the Rht genes (RhtB1a, RhtD1a, RhtB1b, RhtD1b) with structural parameters (JG, PH, CL) (Figure 5).
Figure 5. Correlation analysis of phenotypic parameters and Rht genes
Positive Spearman correlation coefficients are consistently observed among the structural parameters PH, CL, and JG, ranging from R=0.14 to 0.55. Positive correlations with yield are identified for the parameters PH and JG at R=0.14-0.15. As for the genes, a weak positive dependence is noted between the genes RhtB1a and RhtD1b, as well as RhtD1a, with R values ranging from 0.03 to 0.05. The JG index shows a positive correlation with the RhtD1a gene (R=0.14). The PH value exhibits a positive correlation with the RhtD1a and RhtB1b genes, while having a pronounced negative correlation with the other two alleles. Regarding CL, a positive correlation is identified with the RhtD1a gene, aligning with the commonly accepted pattern.
The primary objective of this study was to evaluate the effectiveness of dwarfing genes Rht-B1a/B1b and Rht-D1a/D1b on the phenotype and yield of spring soft wheat under drought conditions. This objective stems from the increasing impact of climate change on agricultural productivity, particularly in arid regions like Kazakhstan. The findings of this study have significant implications for current wheat breeding programs, providing valuable insights into how specific genetic traits can be leveraged to enhance drought resistance and yield. The selection of hybrids through the crossbreeding of promising varieties is a pivotal aspect of wheat breeding. The choice of productive wheat varieties that are well-suited to soil and climatic conditions is very important, it is a need, that heightened by climate change and decreasing yields due to biotic and abiotic stresses. Climate change encompasses not only an increase in air temperature but also a reduction in atmospheric and soil humidity [34]. The continuous reduction in precipitation in Kazakhstan emphasizes the need to accelerate the selection of productive and drought-resistant hybrids and varieties [2].
Despite extensive studies on TY characteristics, the primary focus of global grain crop breeding lies in examining genotype plasticity in adapting to abiotic stresses. Considering all selection parameters in the field is practically unfeasible. In arid climates, yield augmentation is achieved through stem shortening, involving the incorporation of Rht-genes into the breeding process. However, the reduction in PH has a partial impact on yield, particularly noticeable under drought conditions. Wheat PH is a polygenic trait, with about 17 genes identified on 21 chromosomes so far [9], hindering rapid and universal prediction of variety quality and characteristics.
Uncertainty prevails concerning the causes of reduced yield in short-stem varieties; it is observed that dwarf Rht-alleles improve plant productivity by modifying the photosynthesis rate. Several studies indicate a robust correlation between genetic improvements in yield resulting from semi-dwarfism gene introductions and an increase in photosynthesis rate [35]. A recent study highlighted the influence of semi-dwarfism genes on seedling leaf structure, with semi-dwarf plants exhibiting increased stomatal density, thicker leaves, and heightened chlorophyll content [36]. It is essential to acknowledge that early experiments on the effect of semi-dwarfism alleles on photosynthesis were conducted in growth chambers, potentially not reflecting field conditions. Genome-wide association studies (GWAS) on wheat samples from Kazakh, Russian, and foreign selections identified MTAs for PH at locations like the genetic positions corresponding to wheat's Rht-B1, Rht-D1, and Rht8 genes. Notably, germplasm from foreign varieties demonstrated greater breeding potential for yield improvement in Central and Southern regions of the country [37].
The selected parental forms and hybrids in this study exhibited a dependency on dwarfism concerning the presence of the Rht-D1b-gene. In this context, the Rht-B1a gene exhibited a correlation with GL (p<0.06). A significant PH decrease was observed with the presence of the Rht-D1b-gene in the genome, amounting to a reduction of 51.67%. Liu et al. [38] reported that irrigating during the tube insertion stage resulted in a PH increase ranging from 6.60% to 9.70%. In comparison with wild-type alleles Rht-B1a and Rht-D1a, the genetic background of Rht–B1b and Rht-D1b substantially reduced PH by 14.40–15.50% and 16.90-19.10%, respectively. Rht-B1b and Rht-D1b could reduce PH by 29.10–32.50% [38].
However, this gene also resulted in a 30.45% reduction in CL. The presence of the Rht-D1b-gene with a CL greater than 5 cm must be considered when selecting parental forms. Consequently, in 80% of instances, hybrids would exhibit a CL exceeding 4 cm while maintaining a shorter stem.
One of the key factors leading to the predominance of short-statured plants is that those with a PH between 37 cm and 68 cm exhibit resistance to lodging according to the JG index. Plants with a PH exceeding 60 cm exhibit a limit index value of 33 and higher, rendering them more susceptible to lodging.
It was also observed that PH correlates strongly negatively with grain yield during irrigation, while PH correlates positively with the drought coefficient [29].
An augmentation in dry matter content attributed to elongated stems, roots, and increased shoots and leaves can be considered an ancillary outcome that competes with the cultivation of the primary product, grain. This is particularly notable in resource-constrained conditions on arid lands. Photosynthetic activity is linked to final yield [39].
The findings led to the identification of potential productive parent forms such as Saratovskaya 66, Karagandinskaya 22, Xn-10, and Xn-08, as well as hybrids WH134, WH136, WH137, WH145, WH190, and WH192. These selections displayed optimal PH, CL, thereby contributing to enhanced yields in arid climate regions.
Therefore, an examination of the genetic parameters related to dwarfism genes and yield will accelerate the selection of parent forms and hybrids, streamlining the development of stable lines through the integration of both field and laboratory data.
The findings of this study have significant practical implications for wheat breeding programs, particularly those aimed at improving crop performance in arid and semi-arid environments. By prioritizing the selection of genotypes with Rht-B1a/B1b and Rht-D1a/D1b alleles, breeding programs can develop shorter, more drought-resistant wheat varieties. The study highlights the importance of coleoptile length (CL) in drought resilience, recommending the selection of genotypes with longer coleoptiles for better seedling establishment. The integration of field and laboratory data ensures the development of high-yielding, drought-resistant varieties suited to specific climatic conditions. These strategies can lead to substantial yield improvements, as demonstrated by hybrids like WH190, which yielded 2.5 tons per hectare.
The results presented highlight the importance of genetically choosing parent forms and hybrids. However, it is essential to note that additional research is needed to identify a set of genes for more efficient sampling.
This study demonstrates the significant impact of dwarfing genes Rht-B1a/B1b and Rht-D1a/D1b on the phenotype and yield of spring soft wheat under drought conditions. The specific findings reveal that hybrids carrying these alleles exhibit a 15-20% reduction in plant height, which enhances resistance to lodging and optimizes water use.
Alleles of the RhtB1a and RhtD1a genes determine high values of PH, CL, and JG, with the presence of the RhtB1a allele contributing to optimal parameters under drought conditions. Maternal forms represented by varieties of Chinese selection and paternal forms represented by varieties of Kazakh selection are considered the most promising genotypes for hybridization. The presence of even one RhtB1a allele may account for optimal values of structural parameters. Studying wheat genotypes based on dwarfing genes in natural field conditions allows for the identification of an optimal genotype for a specific agro-ecological zone with optimal CL and PH values under drought conditions.
This study was financially supported by the Ministry of Education and Science, Republic of Kazakhstan, within the framework of the Young Scientists Project No. AP13067944 “Molecular SNP-marking of bread wheat by genes TaGW, TaGS, and Rht for grain weight and lodging resistance”.
We would like to express our gratitude to the staff of the Research Platform of Agricultural Biotechnology, as well as to the students of the Faculty of Agronomy of the S.Seifullin Kazakh Agrotechnical Research University.
Table S1: Agronomic and tolerance parameters of parent forms and hybrids; Table S2: Genetic and phenotypic indicators based on stem length; Figure S1: The result of PCR to determine the presence of "green revolution" genes.
Table S1. Agronomic and tolerance parameters of parent forms and hybrids
|
Name of Hybrid |
Parent Forms |
Country |
Plant Height |
Drought Resistance (reference to previous work) |
|
WH 131 |
RAC 875 |
Australia |
Semi-dwarf |
Tolerant |
|
Tauelsizdyk 20 |
Kazakhstan |
High |
Tolerant |
|
|
WH 132 |
Eritrospermum 35 |
Kazakhstan |
High |
Tolerant |
|
Xn-09 |
China |
Semi-dwarf |
Tolerant |
|
|
WH 133 |
Karabalykskaya 25 |
Kazakhstan |
High |
Tolerant |
|
MMF-044 |
Africa |
Semi-dwarf |
Tolerant |
|
|
WH 134 |
Karabalykskaya 25 |
Kazakhstan |
High |
Tolerant |
|
Xn-10 |
China |
Semi-dwarf |
Tolerant |
|
|
WH 135 |
Saratovskaya 66 |
Russian |
High |
Tolerant |
|
Gladius |
Australia |
Semi-dwarf |
Tolerant |
|
|
WH 136 |
Xn-08 |
China |
Semi-dwarf |
Tolerant |
|
Shortandinskaya 2017 |
Kazakhstan |
High |
Tolerant |
|
|
WH 137 |
Xn-10 |
China |
Semi-dwarf |
Tolerant |
|
Astana |
Kazakhstan |
High |
Tolerant |
|
|
WH 138 |
Xn-13 |
China |
Semi-dwarf |
Tolerant |
|
Akmola 2 |
Kazakhstan |
High |
Tolerant |
|
|
WH 139 |
Xn-13 |
China |
Semi-dwarf |
Tolerant |
|
Wyalkatchem |
Australia |
Semi-dwarf |
Tolerant |
|
|
WH 141 |
Xn-03 |
China |
Semi-dwarf |
Tolerant |
|
Akmola 2 |
Kazakhstan |
High |
Tolerant |
|
|
WH 142 |
Xn-03 |
China |
Semi-dwarf |
Tolerant |
|
Aktuybinka |
Kazakhstan |
High |
Tolerant |
|
|
WH 143 |
Xn-03 |
China |
Semi-dwarf |
Tolerant |
|
Lutestsens 141 |
Kazakhstan |
High |
Tolerant |
|
|
WH 145 |
Xn-04 |
China |
Semi-dwarf |
Tolerant |
|
Aktuybinka |
Kazakhstan |
High |
Tolerant |
|
|
WH 146 |
Xn-04 |
China |
Semi-dwarf |
Tolerant |
|
Lutestsens 141 |
Kazakhstan |
High |
Tolerant |
|
|
WH 147 |
Xn-04 |
China |
Semi-dwarf |
Tolerant |
|
Shortandinskaya 2012 |
Kazakhstan |
High |
Tolerant |
|
|
WH 189 |
XN 08 |
China |
Semi-dwarf |
Tolerant |
|
Karagandinskaya 29 |
Kazakhstan |
High |
Tolerant |
|
|
WH 190 |
XN 10 |
China |
Semi-dwarf |
Tolerant |
|
Karagandinskaya 22 |
Kazakhstan |
High |
Tolerant |
|
|
WH 191 |
XN 08 |
China |
Semi-dwarf |
Tolerant |
|
Karabalykskaya 90 |
Kazakhstan |
High |
Tolerant |
|
|
WH 192 |
XN 02 |
China |
Semi-dwarf |
Tolerant |
|
Karagandinskaya 31 |
Kazakhstan |
High |
Tolerant |
|
|
WH 193 |
XN 10 |
China |
Semi-dwarf |
Tolerant |
|
Karagandinskaya 30 |
Kazakhstan |
High |
Tolerant |
Table S2. Genetic and phenotypic indicators based on stem length
|
No. |
Name Hybrid/Parent Form |
Rht B1a |
Rht D1a |
Rht B1b |
RhtD1b |
CL, cm |
PH, cm |
JG, % |
TY, t/ha |
|
1 |
WH 131 |
2 |
0 |
0 |
1 |
4,1±0,05b |
42,02±0.3b |
28,21±0,16b |
1.5±0.1 |
|
2 |
RAC 875 |
0 |
0 |
0 |
1 |
3,72±0,04c |
37.86±0.35c |
19,08±0,3c |
2±0.63 |
|
3 |
Tauelsizdyk 20 |
2 |
2 |
0 |
0 |
5,2±0,06a |
62.72±0,26a |
31,34±0,22a |
1.6±0.1 |
|
4 |
WH 132 |
2 |
0 |
0 |
1 |
4,26±0,08b |
65,2±0,25b |
29,1±0,3b |
1.73±0.3 |
|
5 |
Eritrospermum 35 |
0 |
0 |
1 |
0 |
3,71±0,03c |
67,94±0,3a |
31,61±0,29a |
2.0±0.2 |
|
6 |
Xn-09 |
2 |
2 |
1 |
1 |
5,19±0,06a |
64,74±0,87c |
25,94±0,23c |
2.0±0.2 |
|
7 |
WH 133 |
2 |
2 |
0 |
1 |
5,26±0,07a |
52,62±0,48b |
13,12±0,21c |
1.5±0.1 |
|
8 |
Karabalykskaya 25 |
0 |
2 |
0 |
0 |
4,62±0,04b |
68,42±0,34a |
31,37±0,3b |
2.1±0.2 |
|
9 |
MMF-044 |
0 |
0 |
0 |
1 |
4,18±0,06c |
51,12±0,46b |
33,05±0,22a |
1.7±0.21 |
|
10 |
WH 134 |
2 |
0 |
0 |
1 |
5,24±0,07a |
46,58±0,35b |
24,73±0,23b |
1.6±0.2 |
|
11 |
Karabalykskaya 25 |
0 |
2 |
0 |
0 |
4,62±0,04b |
68,42±0,34a |
31,37±0,3a |
2.1±0.2 |
|
12 |
Xn-10 |
2 |
0 |
0 |
1 |
4,38±0,06c |
37.94±0,52c |
24,15±0,14b |
1.8±0.2 |
|
13 |
WH 135 |
2 |
0 |
0 |
1 |
4,3±0,1b |
53±0,42c |
29,01±0,14b |
1.61±0.1 |
|
14 |
Saratovskaya 66 |
0 |
2 |
0 |
0 |
4,7±0,04a |
67,06±0,41a |
39,06±0,25a |
1.8±0.2 |
|
15 |
Gladius |
0 |
0 |
0 |
1 |
4,32±0,07b |
56,52±0,34b |
24,47±0,22c |
1.9±0.2 |
|
16 |
WH 136 |
2 |
2 |
0 |
0 |
5,26±0,05a |
62,84±0.28b |
28,97±0,3a |
1.6±0.2 |
|
17 |
Xn-08 |
0 |
0 |
1 |
0 |
5,08±0,04a |
65.26±0.32a |
23,7±0,26b |
1.8±0.3 |
|
18 |
Shortandinskaya 2017 |
2 |
2 |
0 |
0 |
4,78±0,06b |
52,98±0,39c |
23,66±0,2b |
1.8±0.3 |
|
19 |
WH 137 |
2 |
0 |
0 |
1 |
5,12±0,06b |
61,02±0,17b |
21,76±0,23c |
1.84±0.1 |
|
20 |
Xn-10 |
0 |
0 |
0 |
1 |
4,44±0,04c |
37.94±0,52c |
24,15±0,14b |
2.1±0.2 |
|
21 |
Astana |
0 |
0 |
0 |
1 |
5,54±0,07a |
66,14±0,2a |
36,05±0,14a |
2.0±0.2 |
|
22 |
WH 138 |
2 |
0 |
1 |
0 |
3,16±0,05c |
43,04±0,29c |
28,67±0,35b |
1.63±0.1 |
|
23 |
Xn-13 |
2 |
0 |
0 |
1 |
4,17±0,04b |
48,08±0,35b |
23,74±0,24c |
1.7±0.3 |
|
24 |
Akmola 2 |
2 |
2 |
1 |
0 |
4,34±0,09a |
68,04±0,23a |
30,23±0,21a |
2.1±0.3 |
|
25 |
WH 139 |
2 |
0 |
0 |
1 |
4,28±0,08a |
38,26±0,31c |
30,85±0,26a |
1.87±0.1 |
|
26 |
Xn-13 |
2 |
0 |
0 |
1 |
4,18±0,03a |
48,08±0,35a |
23,74±0,24b |
2.1±0.3 |
|
27 |
Wyalkatchem |
2 |
2 |
1 |
1 |
3,83±0,04b |
44,88±0,52b |
21,97±0,27c |
1.5±0.15 |
|
28 |
WH 141 |
2 |
0 |
0 |
1 |
3,18±0,06b |
44,02±0,25c |
27,9±0,15b |
1.6±0.1 |
|
29 |
Xn-03 |
2 |
0 |
0 |
1 |
3,18±0,05b |
46,18±0,26b |
25,16±0,16c |
2.1±0.3 |
|
30 |
Akmola 2 |
2 |
2 |
0 |
0 |
4,34±0,09a |
68,04±0,23a |
30,23±0,21a |
1.7±0.3 |
|
31 |
WH 142 |
2 |
0 |
0 |
1 |
4,24±0,09b |
52,48±0,31b |
25,45±0,29b |
1.91±0.2 |
|
32 |
Xn-03 |
2 |
0 |
0 |
1 |
3,18±0,05c |
46,18±0,26c |
25,16±0,16c |
2.1±0.3 |
|
33 |
Aktuybinka |
2 |
2 |
0 |
0 |
5,34±0,07a |
57,94±0,37a |
32,85±0,2a |
1.9±0.3 |
|
34 |
WH 143a |
2 |
0 |
0 |
0 |
4,38±0,07b |
44,66±0,3c |
23,11±0,22c |
1.6±0.2 |
|
35 |
Xn-03 |
2 |
0 |
0 |
1 |
3,18±0,05c |
46,18±0,26b |
25,16±0,16b |
2.1±0.3 |
|
36 |
Lutestsens 141 |
2 |
0 |
0 |
0 |
5,16±0,06a |
58,9±0,5a |
28,94±0,2a |
1.9±0.3 |
|
37 |
WH 145 |
2 |
0 |
0 |
1 |
4,12±0,06b |
47,58±0,23b |
24,62±0,3b |
1.9±0.1 |
|
38 |
Xn-04 |
2 |
0 |
0 |
1 |
4,22±0,04b |
45,18±0,45c |
23,06±0,16c |
1.3±0.3 |
|
39 |
Aktuybinka |
2 |
2 |
0 |
0 |
5,34±0,05a |
57,94±0,37a |
32,85±0,2a |
1.9±0.3 |
|
40 |
WH 146 |
2 |
2 |
1 |
0 |
3,58±0,06c |
42,44±0,35c |
22,07±0,23b |
1.58±0.2 |
|
41 |
Xn-04 |
2 |
0 |
0 |
1 |
4,22±0,04b |
45,18±0,45b |
23,06±0,16c |
1.3±0.3 |
|
42 |
Lutestsens 141 |
2 |
0 |
0 |
0 |
5,16±0,06a |
58,9±0,5a |
28,94±0,2a |
1.9±0.3 |
|
43 |
WH 147 |
2 |
2 |
1 |
1 |
3,54±0,05b |
51,44±0,22b |
27,79±0,22b |
1.42±0.1 |
|
44 |
Xn-04 |
2 |
0 |
0 |
1 |
4,22±0,04a |
45,18±0,45c |
23,06±0,16c |
1.3±0.3 |
|
45 |
Shortandinskaya 2012 |
2 |
0 |
0 |
1 |
4,18±0,04a |
67,24±0,39a |
32,55±0,28a |
1.6±0.3 |
|
46 |
WH189 |
2 |
2 |
0 |
0 |
4,9±0,07c |
61,94±0,14c |
21,81±0,33c |
1.67±0.15 |
|
47 |
XN 08 |
2 |
0 |
0 |
0 |
5,08±0,05a |
64,2±0,33b |
23,7±0,26b |
1.9±0.2 |
|
48 |
Karagandinskaya 29 |
2 |
2 |
0 |
0 |
4,54±0,02b |
75,6±0,24a |
32,79±0,31a |
1.5±0.3 |
|
49 |
WH190 |
2 |
2 |
0 |
0 |
4,42±0,04a |
62.18±0,33b |
23,85±0,18b |
2.5±0.1 |
|
50 |
XN 10 |
2 |
0 |
0 |
1 |
3,49±0,05b |
37.94±0,52c |
24,15±0,14b |
2.1±0.2 |
|
51 |
Karagandinskaya 22 |
2 |
2 |
0 |
0 |
4,46±0,02a |
84,92±0,14a |
33,22±0,19a |
2.1±0.2 |
|
52 |
WH191 |
2 |
0 |
0 |
0 |
5,5±0,05a |
68,86±0,2a |
30,81±0,25a |
1.8±0.2 |
|
53 |
XN 08 |
2 |
0 |
0 |
0 |
5,17±0,05a |
64,2±0,33b |
23,7±0,26c |
1.8±0.3 |
|
54 |
Karabalykskaya 90 |
2 |
0 |
0 |
0 |
4,88±0,04b |
60,76±0,17c |
27,74±0,2b |
1.7±0.3 |
|
55 |
WH192 |
2 |
2 |
0 |
0 |
4,42±0,05a |
64,8±0,27b |
25,77±0,19b |
2.4±0.1 |
|
56 |
XN 02 |
2 |
0 |
0 |
1 |
3,2±0,07b |
47,1±0,25c |
23,03±0,24c |
2.2±0.2 |
|
57 |
Karagandinskaya 31 |
2 |
0 |
0 |
0 |
4,5±0,04a |
68,12±0,24a |
33,03±0,37a |
1.9±0.3 |
|
58 |
WH193 |
2 |
2 |
0 |
1 |
4,4±0,05b |
64,97±0,17b |
26,14±0,24b |
1.85±0.1 |
|
59 |
XN 10 |
2 |
0 |
0 |
1 |
3,49±0,05c |
37.54±0,3c |
24,15±0,14c |
2.1±0.2 |
|
60 |
Karagandinskaya 30 |
2 |
0 |
0 |
0 |
5,24±0,08a |
80.82±0,24a |
33,33±0,22a |
2.0±0.2 |
0- not detected; 1-allele with positive effect; 2-allele with negative effect. a,b,c letters show reliable deviations between parents form and hybrid, the same letters means not reliable differences.
Rht-B1a
Rht-B1b
Rht-D1a
Rht-D1b
Figure S1. The result of PCR to determine the presence of "green revolution" genes
[1] Johnson, E.N., Wang, Z., Geddes, C.M., Coles, K., Hamma, B., Beres, B.L. (2018). Pyroxasulfone is effective for management of bromus spp. in winter wheat in Western Canada. Weed Technology, 32(6): 739-748. https://doi.org/10.1017/wet.2018.70
[2] Karatayev, M., Clarke, M., Salnikov, V., Bekseitova, R., Nizamova, M. (2022). Monitoring climate change, drought conditions and wheat production in Eurasia: The case study of Kazakhstan. Heliyon, 8(1): e08660. https://doi.org/10.1016/j.heliyon.2021.e08660
[3] Bureau of National statistics. Agency for Strategic planning and reforms of the Republic of Kazakhstan. https://stat.gov.kz/en/industries/business-statistics/stat-forrest-village-hunt-fish/publications/, accessed on Apr. 4, 2024.
[4] Crop Prospects and Food Situation – Quarterly Global Report No. 3 September 2022. (2022). FAO, Rome. https://doi.org/10.4060/cc2300en
[5] UNDP Kazakhstan. (2020). Kazakhstan may suffer economic losses in wheat production due to climate change. https://www.undp.org/kazakhstan/stories/kazakhstan-may-suffer-economic-losses-wheat-production-due-climate-change, accessed on Mar. 4, 2024.
[6] Würschum, T., Langer, S.M., Longin, C.F.H., Tucker, M.R., Leiser, W.L. (2017). A modern green revolution gene for reduced height in wheat. The Plant Journal, 92(5): 892-903. https://doi.org/10.1111/tpj.13726
[7] Chandler, P.M., Harding, C.A. (2013). ‘Overgrowth’ mutants in barley and wheat: New alleles and phenotypes of the ‘Green Revolution’ DELLA gene. Journal of Experimental Botany, 64(6): 1603-1613. https://doi.org/10.1093/jxb/Fert022
[8] Ellis, M., Spielmeyer, W., Gale, K., Rebetzke, G., Richards, R. (2002). "Perfect" markers for the Rht-B1b and Rht-D1b dwarfing genes in wheat. Theoretical and Applied Genetics, 105(6-7): 1038-1042. https://doi.org/10.1007/s00122-002-1048-4
[9] Zhang, W., Li, H., Zhi, L., Su, Q., Liu, J., Ren, X., Meng, D., Zhang, N., Ji, J., Zhang, X., Li, J. (2020). Functional markers developed from TaGS3, a negative regulator of grain weight and size, for marker-assisted selection in wheat. The Crop Journal, 8(6): 943-952. https://doi.org/10.1016/j.cj.2020.03.003
[10] Ukozehasi, C., Ober, E.S., Griffiths, H. (2022). The other mechanisms by which the RHT genes improve the harvest index of wheat. Plants, 11(21): 2837. https://doi.org/10.3390/plants11212837
[11] Voronov, S., Pleskachiov, Y., Shitikova, A., Zargar, M., Abdelkader, M. (2023). Diversity of the biological and proteinogenic characteristics of quinoa genotypes as a multi-purpose. Crop. Agronomy, 13(2): 279. https://doi.org/10.3390/agronomy13020279
[12] Anzooman, M., Dang, Y.P., Christopher, J., Mumford, M.H., Menzies, N.W., Kopittke, P.M. (2018). Greater emergence force and hypocotyl cross sectional area may improve wheat seedling emergence in sodic conditions. Plant Science, 277: 188-195. https://doi.org/10.1016/j.plantsci.2018.09.007
[13] Rebetzke, G.J., Ellis, M.H., Bonnett, D.G., Richards, R.A. (2007). Molecular mapping of genes for coleoptile growth in bread wheat (Triticum aestivum L.). Theoretical and Applied Genetics, 114: 1173-1183. https://doi.org/10.1007/s00122-007-0509-1
[14] Xu, D., Hao, Q., Yang, T., Lv, X., Qin, H., Wang, Y., Jia, C., Liu, W., Dai, X., Zeng, J., Zhang, H., He, Z., Xia, X., Cao, S., Ma, W. (2023). Impact of “Green Revolution” gene Rht-B1b on coleoptile length of wheat. Frontiers in Plant Science, 14: 1147019. https://doi.org/10.3389/fpls.2023.1147019
[15] Li, P., Chen, J., Wu, Y., Zhang, J., Chu, C., See, D., Brown-Guedira, G., Zemetra, R., Souza, E. (2011). Quantitative trait loci analysis for the effect of Rht-B1 dwarfing gene on coleoptile length and seedling root length and number of bread wheat. Crop Science, 51(6): 2561-2568. https://doi.org/10.2135/cropsci2011.03.0116
[16] Guan. P., Shen, X., Mu, Q., Wang, Y., Wang, X., Chen, Y., Zhao, Y., Chen, X., Zhao, A., Mao, W., Guo, Y., Xin, M., Hu, Z., Yao, Y., Ni. Z., Sun, Q., Peng, H. (2020). Dissection and validation of a QTL cluster linked to Rht-B1 locus controlling grain weight in common wheat (Triticum aestivum L.) using near-isogenic lines. Theoretical and Applied Genetics, 133(9): 2639-2653. https://doi.org/10.1007/s00122-020-03622-z
[17] Zotova, L.P., Kipshakbayeva, G.A., Tleulina, Z.T. (2019). Vegetatsionnyy period urozhaynost' yarovoy myagkoy pshenitsy v usloviyakh Akmolinskoy oblasti Severnogo Kazakhstana [The growing season and yield of spring soft wheat in the conditions of the Akmola region of Northern Kazakhstan]. In Collection of Materials of the International Scientific-Practical Conference Devoted to the 85th Anniversary of the Kazakh Research Institute of Agriculture and Plant Industry, Almaty, pp. 192-197.
[18] Shavrukov, Y., Zhumalin, A., Serikbay, D., Botayeva, M., Otemisova, A., Absattarova, A., Sereda, G., Sereda, S., Shvidchenko, V., Turbekova, A., Jatayev, S., Lopato, S., Soole, K., Langridge, P. (2016). Expression level of the DREB2-type gene, identified with amplifluor SNP markers, correlates with performance, and toleranceto dehydration in bread wheat cultivars from Northern Kazakhstan. Frontiers in Plant Science, 7: 1736. https://doi.org/10.3389/fpls.2016.01736
[19] Borlaug, N.E. (1968). Wheat breeding and its impact on world food supply. In Proceedings of the 3rd International Wheat Genetics Symposium, Canberra, Butterworths, Sydney, pp. 1-36.
[20] Yapiyev, V., Gilman, C., Kabdullayeva, T., Suleimenova, A., Shagadatova, A., Duisembay, A., Naizabekov, S., Mussurova, S., Sydykova, K., Raimkulov, I., Kabimoldayev, I., Abdrakhmanova, A., Omarkulova, S., Nurmukhambetov, D., Kudarova, A., Malgazhdar, D., Schönbach, C., Inglezakis, V. (2018). Top soil physical and chemical properties in Kazakhstan across a north-south gradient. Scientific Data, 5: 180242. https://doi.org/10.1038/sdata.2018.242
[21] Selyaninov, G.T. (1928). About climate agricultural estimation. In Proceedings on Agricultural Meteorology, 20: 165-177.
[22] National Hydrometeorological Service of Kazakhstan - Kazhydromet. https://www.kazhydromet.kz/ru/weather/in_city/20/347, accessed on Apr. 13, 2024.
[23] Tao, H., Feng, H., Xu, L., Miao, M., Yang, G., Yang, X., Fan, L. (2020). Estimation of the yield and plant height of winter wheat using UAV-based hyperspectral images. Sensors, 20(4): 1231. https://doi.org/10.3390/s20041231
[24] Sidhu, J.S., Singh, D., Gill, H.S., Brar, N.K., Qiu, Y., Halder, J., Tameemi, R.A., Turnipseed, B., Sehgal, S.K. (2020). Genome-wide association study uncovers novel genomic regions associated with coleoptile length in hard winter wheat. Frontiers in Genetics, 10: 1345. https://doi.org/10.3389/fgene.2019.01345
[25] Reimova, B.T., Kudaibergenova, K.E. (2022). Selection of breeding samples of winter barley in the conditions of the Republic of Karakalpakstan. Theory and Practive of Contemporary Science, 11(89): 120-124.
[26] Aboul-Maaty, N.A.F., Oraby, H.A.S. (2019). Extraction of high-quality genomic DNA from different plant orders applying a modified CTAB-based method. Bulletein of the National Research Centre, 43: 25. https://doi.org/10.1186/s42269-019-0066-1
[27] Zotova, L.P., Sereda, T.G., Gadzhimuradova, A.M., Liang, C., Zhirnova, I.A., Ilyasova, D.Z. (2023). Early inheritance of the short-stem and coleoptile length trait by hybrids between foreign and Kazakh varieties in the conditions of central and northern Kazakhstan. Bulletin of Science of the Kazakh Agrotechnical Research University named after S. Seifullin, 1(116): 21-33. https://doi.org/10.51452/kazatu.2023.1(116).1286
[28] Islam, M., Obour, A., Malay, S., Nachtman, J., Wendy, C., Baumgartner, R. (2013). Grain yield, forage yield, and nutritive value of dual-purpose small grains in the Central High Plains of the USA. Crop Management, 12(1): 1-8. https://doi.org/10.1094/CM-2012-0154-RS
[29] Gao, Z., Wang, Y., Tian, G., Zhao, Y., Li, C., Cao, Q., Han, R., Shi, Z., He, M. (2020). Plant height and its relationship with yield in wheat under different irrigation regime. Irrigation Science, 38(4): 365-371. https://doi.org/10.1007/s00271-020-00678-z
[30] Pumpa, J., Martin, P., McRae, F., Coombes, N. (2013). Coleoptile length of wheat varieties. NSW Government. Department of Primary Industries.
[31] Rebetzke, G., Fletcher, A., Micin, S., (CSIRO Agriculture and Food) and Callum Wesley, (Charlesville Ag, Southern Cross WA). (2021). On-farm assessment of new long-coleoptile wheat genetics for improving grain yield with deep sowing. Australian Government, Grains Research and Development Corporation.
[32] Ivanova, Y.S., Fomina, M.N. (2017). Morphological parameters of stem lodging resistance in accessions of hulless oats collection. Agricultural Science Euro-North-East, (3): 15-21.
[33] Jaenisch, B.R., Munaro, L.B., Jagadish, S.V.K., Lollato, R.P. (2022). Modulation of wheat yield components in response to management intensification to reduce yield gaps. Frontiers in Plant Science, 13: 772232 https://doi.org/10.3389/fpls.2022.772232
[34] Elahi, I., Saeed, U., Wadood, A., Abbas, A., Nawaz, H., Jabbar, S. (2022). Effect of climate change on wheat productivity. In: Ansari, M.-ur-R. (ed) Wheat - Recent Advances, 23-35. https://doi.org/10.5772/intechopen.103780
[35] Evans, J.R. (2013). Improving photosynthesis. Plant Physiology, 162(4): 1780-1793. https://doi.org/10.1104/pp.113.219006
[36] Nenova, V.R., Kocheva, K.V., Petrov, P.I., Georgiev, G.I., Karceva, T.V., Borner, A. (2014). Wheat Rht-B1 dwarfs exhibit better photosynthetic response to water deficit at seedling stage compared to the wild type. Journal of Agronomy and Crop Science, 200(6): 434-443. https://doi.org/10.1111/jac.12090
[37] Turuspekov, Y., Baibulatova, A., Yermekbayev, K., Tokhetova, L., Chudinov, V., Sereda, G., Ganal, M., Griffiths, S., Abugalieva, S. (2017). GWAS for plant growth stages and yield components in spring wheat (Triticum aestivum L.) harvested in three regions of Kazakhstan. BMC Plant Biology, 17: 190. https://doi.org/10.1186/s12870-017-1131-2
[38] Liu, Y., Zhang, J., H, Y., Chen, J. (2017). Dwarfing genes Rht4 and Rht-B1b affect plant height and key agronomic traits in common wheat under two water regimes. Field Crops Research, 204: 242-248. https://doi.org/10.1016/j.fcr.2017.01.020
[39] Brown, M.M., Martin, J.M., Jobson, E.M., Hogg, A.C., Carr, P.M., Giroux, M.J. (2021). Evaluating the impact of Rht hypomorphic mutations in durum wheat. Crop Science, 62(1): 247-258. https://doi.org/10.1002/csc2.20672