Reza Muhammad Riady![]() | Ren Fitriadi
| Ren Fitriadi![]() | Kasprijo
| Kasprijo![]() | Dini Ryandini
| Dini Ryandini![]() | Mustika Palupi*
| Mustika Palupi*![]() | Purnama Sukardi
| Purnama Sukardi![]() | Mohammad Nurhafid
| Mohammad Nurhafid![]() | Ahmad Musa
| Ahmad Musa![]() | Ruzkiah Asaf
| Ruzkiah Asaf![]()
© 2024 The authors. This article is published by IIETA and is licensed under the CC BY 4.0 license (http://creativecommons.org/licenses/by/4.0/).
OPEN ACCESS
Activities in the Minapadi fish farming ecosystem are closely related to the role of microorganisms, including degrading bacteria found in sediment. Therefore, in the short term, this research aims to determine the proportion and activity index of proteolytic bacteria isolated from the sediment of the mina rice cultivation system. Research is expected to find the best isolate as a probiotic agent which can be used in fish cultivation in the long run. The method used is observation. Sediment sampling was carried out using a purposive random sampling method in two ponds of the Minapadi fish cultivation system which have different characteristics. Sampling was taken at Block A and Block E, at each location 3 points were taken, namely inlet, middle and outlet. The results of the study showed that the proportion of proteolytic bacteria was found in the sediment in varying proportions in Block A and Block B with respective values, namely inlet (72%; 68%), middle (76%; 72%), and outlet (52%; 48%). Proteolytic activity index with successive values: inlet (0.4-4.0: 0.2-2.5), middle 0.6-6.0: 0.4-4.0 and outlet (0.3-3.8: 0.6-4.0). Five bacterial isolates were found to have the highest proteolytic activity index (>4), namely SAT.2, SAT.7, SAT.12, SAT.11, and SET.10. This research obtained interesting results regarding the activity index and proportion of proteolytic bacteria isolated from rice fish sediments. Most of the proportions of proteolytic bacteria from both blocks showed higher proportions than the general bacteria obtained. Therefore, it can be concluded that bacterial isolates are capable of biodegrading proteins found in organic materials.
biodegradation, proteolytic bacteria, activity index, rice-fish farming system, sediments
Rice-fish farming system is an integrated method that combines fish and rice cultivation in one location [1]. This system is highly effective and efficient in rice fields with sufficient water availability throughout the year, and the dynamic interaction associated forms a unique ecosystem. Fish activity in ponds acts as a biocontrol agent in reducing pest and weed attacks on rice plants [2]. Meanwhile, rice plants provide a protective shelter for fish during high water temperatures, aiding in the reduction of ammonia concentrations in water, and enhancing nitrogen solubility in sediments [3]. A rice-fish farming system has a positive effect on the farmer's economy as well because it creates an ecosystem by cultivating fish using rice-fish farming techniques, which can enhance yield diversity, improve soil and water fertility, reduce the use of chemical fertilizers, limit the growth of other plants, and reduce pests and diseases [4]. The advantages of the rice-fish farming ecosystem are influenced by various things, especially the impact of environmental interactions, organisms, and microorganisms found in the Minapadi pond environment. Considering these advantages, cultivation with the system has potentials for further development and study. In the rice-fish farming cultivation system, microorganisms, especially bacteria, have an important role in increasing the productivity of fish and rice cultivation [5].
The new ecosystem created through rice-fish farming system is closely related to the role of microorganisms, including bacteria present in the environment [5]. However, the potential of these bacteria has not been fully explored. One of their functions is the degradation of organic matter accumulated in sediments [6, 7]. Organic matter contained in rice-fish farming system sediments mostly originates from leftover feed, fish feces, as well as remains of rice plants that fall and accumulate at the bottom of the pond [8, 9]. According to MacKay [8] fish cultivation using the rice-fish farming system increases the accumulation of organic matter by 0.09%, total nitrogen by 0.04%, total phosphorus by 0.38%, nitrogen availability by 22 ppm, and phosphorus availability by 2 ppm. These materials contain undegraded residual proteins which affect water quality [10], hence, there is a need to explore and screen for bacteria capable of breaking down organic matter present in rice-fish farming system sediments [11]. One group of microorganisms that play a role in protein degradation is proteolytic bacteria [12]. In the environment, proteolytic bacteria will break down organic compounds from the protein group so that they do not accumulate as ammonia which can pollute the aquatic environment [13]. In another study, proteolytic bacteria have a positive correlation and both play a role in bioremediation [14, 15].
Proteolytic bacteria produce protease enzymes [16], which serve to hydrolyze proteins into amino acids [17]. Protease-producing bacteria can improve protein digestibility and host growth, as well as reduce organic pollutants in aquaculture [5, 18]. The proteolytic bacteria have a unique catalytic mechanism and are therefore well-suited to bioremediation due to their broad substrate specificity and high resistance. Some examples of these bacteria include the genus Pseudomonas, Vibrio, Bacillus [19, 20], Pseudoalteromonas [18], Alcaligenes, Flavobacterium, Corynebacterium [21], Staphylococcus, and Plesiomonas [16]. Proteolytic bacteria can be detected by measuring the hydrolysis zones around colonies on selective growth media that contain skim milk. The media used for proteolytic bacteria is designed to contain 80% casein, which is a type of protein. The hydrolysis process breaks down protein molecules by breaking the peptide chains in the medium. Proteolytic activity is measured by dividing the diameter of the hydrolysis area by the diameter of the bacterial colony, to obtain a result called the activity index [22-24]. Different bacteria have varying proteolytic activity index, for example, Hastuti et al. [20] obtained values ranging from 1.62-2.94, while Mulyasari et al. [25] recorded the largest index of 3.3. The higher proteolytic activity index, the greater the ability of bacteria to produce protease enzymes.
The presence of proteolytic bacteria in the aquatic environment is generally higher than other bacteria. This is closely related to the role of proteolytic bacteria. In rice cultivation environments, bacteria play a crucial role in fish, rice plants, and environmental media. Proteolytic bacteria play a crucial role in accelerating the metabolic breakdown of feed compounds in fish so that they can be absorbed rapidly. In rice plants, proteolytic bacteria play a role in the process of degradation and absorption of nutrients by plant roots. Apart from that, proteolytic bacteria can synergize with the nitrifying bacteria group in mineralizing organic materials in the nitrogen cycle. In the environment, proteolytic bacteria have a role in degrading protein compounds so that they do not become environmental pollutants such as ammonia. This research seeks to discuss more complexly the role of proteolytic bacteria in the integrated environment of fish and rice plants, namely mina padi. Additionally, the development of mina padi fish cultivation in Indonesia is progressing very rapidly, so probiotics will be needed in the future in order to support the growth of the plant.
2.1 Research time and place
The research was conducted between September 10 to November 10, 2022, while the data processing and thesis preparation continued from January to June 2023. Sediments sampling was carried out in rice-fish farming system pond in Panembangan Village, Cilongok District, Banyumas Regency. The preparation of tools and materials for conducting the Gram test, catalase, and oxidase tests of proteolytic bacteria was performed at the Microbiology Laboratory of Muhammadiyah Purwokerto University, and the Research Laboratory, Faculty of Fisheries and Marine Science, Jenderal Soedirman University.
2.2 Sample collection
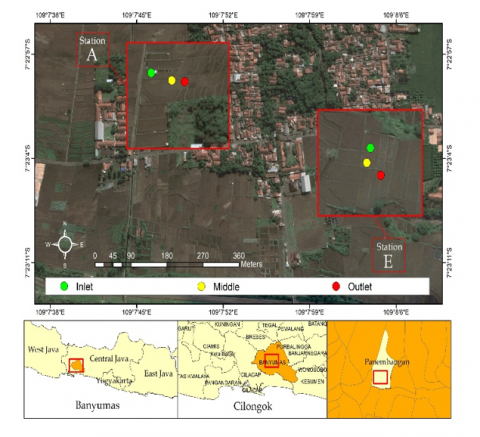
Sediment sampling was carried out using a purposive random sampling method in two rice fish farming system ponds which have different characteristics. The first is Block A, at this location 3 points were taken, namely inlet, middle and outlet. This location is characterized by sandy sediments that are fed by water directly from the mountains. Then second is Block E, at this location three points were taken, namely inlet, middle and outlet. This location has a muddy sediment type and is irrigated from water passing through residential areas (Figure 1). The sampling process was carried out using a sterile spoon, where sediments were taken at the top layer of the surface, placed in a petri dish under sterile conditions, and stored in a cool box for further processing in the laboratory.
Figure 1. Sediments sampling locations of rice-fish farming system ponds
2.3 Bacteria isolation
Bacteria isolates were taken by weighing 0.1 g of sediments samples, which were suspended in 1 mL of physiological solution (NaCl 0.8%), and homogenized with a vortex. The sample suspension was serially diluted three (10-1-10-3). For the 10-1 dilution, 0.5 mL of the sample suspension was taken and homogenized with 4.5 mL of physiological solution in the first tube. And then, take 0.5ml of the tube (10-1) dilution and add in to the tube (10-2) dilution than homogenies. This process was repeated for the subsequent tubes creating dilutions 10-3. Subsequently, 0.5 mL of each dilution sample was taken and cultured using the pour plate method on TSA media. Incubation was carried out for 18-24 hours at 28℃ to facilitate bacteria growth [26].
2.4 Bacteria abundance calculation
Colonies that grew on each TSA media, namely at dilutions 10-1 to 10-3, were counted entirely using a counter tool. Afterward, the abundance of sediments bacteria was calculated using the total plate count (TPC) method with colonies numbering 30-300 [27]. This calculation referred to the equation outlined by Madigan and Martinko [28].
${Number\, of\, bacteria}\left(\frac{C F U}{g}\right)={Number\, of \,colony}\times \frac{1}{ Dilution} \times \frac{1}{Culture volume} \times \frac{1}{Sample weight}$
2.5 Observation of bacteria morphology
Bacteria that grew on TSA media were first observed macroscopically for colony morphology, comprising color, shape, elevation, edge, and size [29]. A total of 25 isolates of bacteria colonies that grew separately were taken from each sample dilution. The selection was based on visible morphological differences, and bacteria were stocked using the streak plate technique on TSA media.
2.6 Proteolytic activity testing
Proteolytic activity was evaluated by transferring bacteria culture using an ose needle into TSA media, followed by streaking on skim milk agar media, and incubation for 48 hours at 28℃. Bacteria isolates with proteolytic activity showed a clear zone around the colony. The diameter of both bacteria colony and the clear zone formed was measured to determine proteolytic activity index [30, 31]. The measurement results were entered into proteolytic activity calculation formula as quantitative data [26]. The formula for calculating proteolytic activity index referred to Firliani et al. [32]:
${Proteolytic\, Activity} =\frac{{ Clear\, zone\, total\, diameter }- { bacterial\, colony \,diameter }}{{ bacterial\, colony \,diameter }}$
Proportion of proteolytic bacteria was calculated using the formula by Sinatryani [33]:
${Proportion\, of\, proteolytic\, bacteria}=\frac{ { Number\, of\, proteolytic\, bacteria\, colonies \,obtained }}{{ Total \,number\, of\, colonies\, observed }} \times 100$
2.7 Gram observation
For the gram observation, 1 drop of 3% KOH was dripped on the glass object, then a single bacteria isolate was taken from each culture stock (1 ose) and placed on the stock. Positive Gram result was characterized by no mucus formation when the ose was lifted, while negative Gram outcome was indicated by the formation of mucus [34].
2.8 Catalase test
Catalase testing was performed using one drop of hydrogen peroxide (H2O2) solution dripped on the glass object. Proteolytic bacteria isolates were taken up to 1 ose from each culture stock and then observed on a glass object containing H2O2 solution. Positive results were characterized by the formation of gas bubbles, with the appearance of several bubbles indicating a weak reaction. Meanwhile, negative results were characterized by the absence of gas bubble formation [35].
2.9 Oxidase test
The oxidase test involved taking 1 ose of proteolytic bacteria isolate, followed by observation on a glass object added with 1-2 drops of tetramethyl-blue reagent and covered with filter paper. Positive oxidase result was marked by a change in the color of the paper to blue or dark purple, while the absence of color change indicated a negative result [36].
2.10 Data analysis
The data obtained on bacteria abundance, colony morphology, proteolytic activity index, bacteria abundance, Gram percentage, catalase, and oxidase test analyzed statistics and were presented in the form of figures, tables, and graphs. Therefore, the data were analyzed descriptively and compared with the literature.
3.1 Bacteria abundance in rice-fish farming system sediments
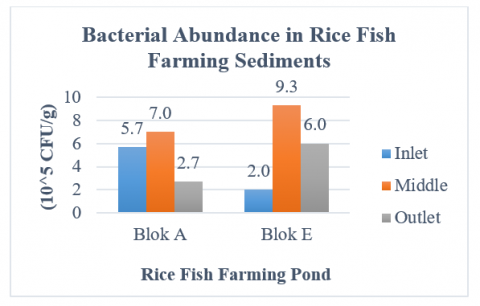
The abundance of bacteria found in rice-fish farming system sediments showed varied values at each sampling point, with greater abundance in the middle point compared to the inlet and outlet, as shown in Figure 2.
Figure 2. Bacteria abundance in rice-fish farming system sediments
Based on Figure 2, the bacterial abundance value in block A at the inlet point is 5.7 × 105 CFU/g, the middle point is 7.0 × 105 CFU/g, and the outlet point is 7.0 × 105 CFU/g. Meanwhile, the bacterial abundance value in block E at the inlet point was 2.0 × 105 CFU/g, the middle point was 9.3 × 105 CFU/g, and the outlet point was 6.0 × 105 CFU/g. The results of the study showed quite high values compared to research by Kasprijo et al. [37] which reported an abundance of bacteria from Minapadi pond sediment of 1.8 × 105 CFU/g. The high value of bacterial abundance found in pond sediments is an indicator of organic material pollution in the pond [38]. Differences in abundance values at each point indicate the presence of different organic materials.
Organic matter contained in rice-fish farming system sediments accumulated at the middle point due to water flow patterns, leading to increased bacteria growth in the center of the pond. There were also significant differences in the flow of water. In block A, the water directly originated from the mountains, while in block E, it passed through the community settlement before entering the pond. This disparity affected the accumulation of organic matter contained in sediments. Water flow that has passed through community settlements may carry organic matter from household waste. The accumulation of organic matter caused an increase in the abundance of bacteria within the sediments [9]. The higher the organic matter in the pond, the greater bacteria abundance [39].
Another factor that affected the difference in the abundance of bacteria was the type of sediments. In Block A rice-fish farming system pond, the sediments were majorly sandy, while Block E had a muddy type. These differences significantly affected the abundance and community of bacteria [40]. According to Irene et al. [41], the presence of bacteria was more predominant in mangrove areas with sandy and muddy sediments types.
3.2 Morphology of bacteria in rice-fish farming system sediments
The morphology of bacteria colonies observed varied significantly, consisting of 102 different types, out of the 150 isolates obtained. The morphological variations were classified based on Bergey's Manual of Determinative Bacteriology [29] which included shape, elevation, edge, color, and size. Based on the results, the colony shapes obtained were circular, irregular, filamentous, rhizoid, spindle, and punctiform, while elevations consisted of convex, crateriform, raised, pulvinate, umbonate, and flat. Furthermore, edges comprised entire, undulate, lobate, and filamentous. The colony colors included milky, clear, grayish, creamy, brownish, and yellowish-white, yellow, white, brownish-yellow, creamy brown, and light yellow, while sizes consisted of small, medium, and large.
The differences in colony morphology indicated that bacteria isolates stemmed from various species [42]. Factors that affect variations in shape, color, and general morphological characteristics of bacteria include age, certain growth requirements, environment (biotic and abiotic), food (growth medium), temperature, pH, and free oxygen [43, 44]. According to Zuraidah et al. [45], the environmental conditions of bacteria life may cause changes in the morphological and physiological properties.
3.3 Proteolytic bacteria proportion in rice-fish farming system sediments
Proportion of proteolytic bacteria was indicated by the presence of protease enzyme activity produced. This was showed by the formation of a clear zone around bacteria colonies (Figure 3). The size of the clear zone indicates activity of protease enzymes.
Figure 3. Protease enzyme activity produced by proteolytic bacteria
The clear zone that forms around bacterial colonies is the result of breaking down protein peptide bonds into simpler peptides [46]. Casein contained in skim milk is a protein consisting of phosphoprotein which binds to calcium and forms calcium calceinate salt which makes the growth medium for proteolytic bacteria white [23, 47]. Protease enzymes produced by bacteria cause casein to hydrolyze into soluble amino acids so that the white color in the growth medium disappears and a clear zone form around the bacterial colony [48]. The proportion of bacteria capable of producing protease enzyme activity was calculated to determine the distribution of proteolytic bacteria found in Rice-Fish farming sediments. According to Rice-Fish systems sediments, proteolytic bacteria play an important role in the degradation of proteins found in organic matter.
Proteolytic activity test showed that among the 150 bacteria isolates obtained from rice-fish farming system sediments, 97 exhibited proteolytic activity. The highest proportion of proteolytic bacteria was found at the middle point, followed by the inlet and outlet, as presented in Table 1.
Table 1. Proteolytic bacteria proportion in rice-fish farming system sediments
|
Sample |
Number of Isolates |
Number of Proteolytic Isolates |
Proportion (%) |
|||
|
Block A |
Block E |
Block A |
Blok E |
Block A |
Block E |
|
|
Inlet |
25 |
25 |
18 |
17 |
72 |
68 |
|
Middle |
25 |
25 |
19 |
18 |
76 |
72 |
|
Outlet |
25 |
25 |
13 |
12 |
52 |
48 |
Based on Table 1, the highest proportion of proteolytic bacteria was found at the middle point in both blocks, namely 76% in block A and 72% in block E. The accumulation of protein-containing organic matter caused the high proportion of proteolytic bacteria found at the midpoint of rice-fish farming system pond sediments. This organic matter stemmed from leftover feed and fish feces [11], as well as rice plant parts including roots, straw, and flowers present in the pond. These materials serve as a source of protein for proteolytic bacteria [8]. According to previous research, the abundance of protein sources in the pond facilitates the dominance of proteolytic bacteria to carry out their function as protein decomposers [49]. The high protein content provides a suitable environment for their growth and development [50].
3.4 Proteolytic activity index of bacteria in rice-fish farming system sediments
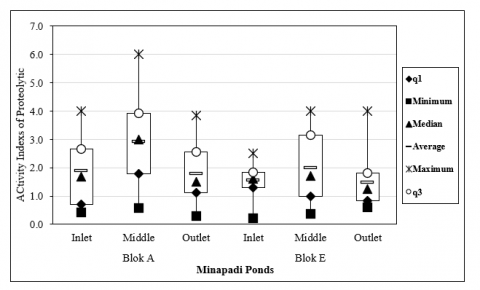
Proteolytic bacteria activity index was obtained by dividing the diameter of the clear zone by that of the colony. The highest average proteolytic activity index value was obtained at 48 hours of incubation time, as presented in Figure 4.
Figure 4. Proteolytic activity index of bacteria in rice-fish farming system sediments
Description: Quartile 1 (q1): The lowest distribution of proteolytic index data; Minimum: The lowest proteolytic index value; Median: The middle value of proteolytic index; Mean: Average value of proteolytic index; Maximum: Highest proteolytic index value; Quartile 3 (q3): Highest proteolytic index data distribution; Box plot: Proteolytic index data set.
The proteolytic index values obtained in our research ranged from 0.1 - 6.0. The distribution of proteolytic bacteria obtained from each sampling point is presented in the box plot in Figure 4. There are differences in the proteolytic index at each sampling point. These differences indicate the ability of bacteria to produce protease enzymes [51], as also stated by Paskandani et al. [52]. Each bacteria species has varying capabilities of producing protease enzymes with different activity index [16]. Based on the results, certain bacteria isolates did not yield proteolytic index, evidenced by the absence of clear zones around colonies due to the inability to produce protease enzymes [53]. Another factor that affected the difference in proteolytic index was the length of incubation time. This factor influenced the production of protease enzymes [54].
Based on the results, bacteria isolates obtained had a high proteolytic activity index > 3, characterized by the formation of large clear zones [55]. According to Irene et al. [41], proteolytic index was categorized into 4 groups, namely very low (≤ 1), low (1-2), medium (2-3), and high (> 3). Proteolytic activity index values obtained are presented in Table 2. Of the 97 bacterial isolates that have proteolytic activity index, then narrowed down to 25 bacterial isolates with the highest proteolytic activity index of more than three (>3). The high index of proteolytic activity indicates the ability of bacteria to produce protease enzymes. The amount of protease enzyme produced shows the potential of bacteria as environmental bioremediation agents [56].
Table 2. Highest proteolytic bacteria activity index
|
Block |
Point |
Best Isolate |
Proteolytic Activity Index (cm) |
|
Block A |
Inlet |
SAI.8 |
4.0 |
|
SAI.15 |
3.8 |
||
|
SAI.16 |
3.7 |
||
|
SAI.23 |
3.8 |
||
|
Middle |
SAT.1 |
3.1 |
|
|
SAT.2 |
4.2 |
||
|
SAT.6 |
3.6 |
||
|
SAT.7 |
6.0 |
||
|
SAT.11 |
4.2 |
||
|
SAT.12 |
5.5 |
||
|
SAT.13 |
4.0 |
||
|
SAT.14 |
3.7 |
||
|
SAT.21 |
3.4 |
||
|
Outlet |
SAO.2 |
3.8 |
|
|
SAO.7 |
3.3 |
||
|
SAO.15 |
3.3 |
||
|
Block E |
Inlet |
SEI.25 |
3.4 |
|
Middle |
SET.1 |
4.0 |
|
|
SET.2 |
4.0 |
||
|
SET.10 |
4.2 |
||
|
SET.20 |
4.0 |
||
|
SET.21 |
3.8 |
||
|
SET.22 |
3.7 |
||
|
SET.23 |
3.5 |
||
|
Outlet |
SEO.5 |
4.0 |
Description: SAI.8: Sediments block A inlet isolate 8, etc.; SAT.1: Sediments block A middle isolate 1, etc; SAO.2: Sediments block A outlet isolate 2, etc; SEI.25: Sediments block E inlet isolate 25; SET.1: Sediments block E middle isolate 1, etc; SEO. 5: Sediments block E outlet isolate 5.
This study obtained interesting results regarding the activity index and proportion of proteolytic bacteria isolated from rice-fish sediments. Most of the proportions of proteolytic bacteria from both blocks showed higher proportions than the general bacteria obtained. Then as many as 25 bacterial isolates showed an activity index in the high category. Several studies state that the presence and high activity index of proteolytic bacteria shows a positive correlation with the protein content in the environment. The higher the number of proteolytic bacteria, the higher the protein content in the environment. This is because protein is a carbon source for proteolytic bacteria so that the presence and activity of proteolytic bacteria are closely related [57]. The results of this research can illustrate that the protein content in the rice-fish system environment has organic material, especially high protein. The presence of proteolytic bacteria in this environment has a very good impact on the environment. The buildup of protein in the environment will cause increased levels of ammonia which can be toxic to cultivated biota. However, the presence of proteolytic bacteria will break down protein compounds and synergize with nitrifying bacteria which will convert them into nitrites and nitrates so that they can be beneficial for the environment [15, 58]. The potential and abilities of these bacteria continue to be developed by researchers in order to obtain environmentally friendly biotechnology products as bioremediation agents.
3.5 Gram test results, catalase and oxidase activity of proteolytic bacteria
The gram, catalase, and oxidase tests were carried out to further assess the characteristics of proteolytic bacteria, and the results obtained are presented in Table 3.
Among the 97 bacteria isolates with proteolytic activity, 57 were Gram-positive, while 40 were negative. These results show the predominance of Gram-positive proteolytic bacteria compared to the negative ones. Gram-positive bacteria are generally found in sediments and soils [59], have thick peptidoglycan walls that protect cells from extreme environmental conditions, and also contribute to the degradation of complex compounds [60].
Based on the results, 77 isolates were found to have positive catalase activity, indicating that the majority of proteolytic bacteria produced the catalase enzyme. This enzyme is used by bacteria to catalyze the decomposition of hydrogen peroxide into water and oxygen, as well as methanol, ethanol, formic acid, thiols, and phenols [61]. Hydrogen peroxide is toxic to bacteria cells because it potentially inactivates enzymes in cells, causing harm [62]. The elevated production of catalase enzymes was caused by increased oxygen levels, the large accumulation of H2SO2 in cells, and high organic matter. The prevalence of catalase-producing bacteria can serve as an indicator of pollution levels [63, 64].
A total of 59 proteolytic bacteria isolates were found to have positive oxidase activity, indicating that the majority of proteolytic bacteria produced the enzyme, cytochrome oxidase [65]. This enzyme plays a crucial role in the electron transport chain during aerobic respiration, enabling bacteria to reduce molecular oxygen, leading to the production of water and energy as byproducts. Bacteria lacking this enzyme rely on fermentation rather than respiration for energy metabolism [66-68]. The production of cytochrome oxidase enzyme can also be used to identify certain bacteria [69].
Table 3. Gram, catalase, and oxidase test results of proteolytic bacteria
|
Block |
Point |
Gram Test |
Catalase Test |
Oxidase Test |
|||
|
(+) |
(-) |
(+) |
(-) |
(+) |
(-) |
||
|
Block A |
Inlet |
6 |
12 |
16 |
2 |
7 |
11 |
|
Middle |
14 |
5 |
13 |
6 |
8 |
11 |
|
|
Outlet |
6 |
7 |
10 |
3 |
7 |
6 |
|
|
Block E |
Inlet |
10 |
7 |
11 |
6 |
14 |
3 |
|
Middle |
17 |
1 |
17 |
1 |
15 |
3 |
|
|
Outlet |
4 |
8 |
10 |
2 |
8 |
4 |
|
Table 4. Characteristics of proteolytic bacteria isolates with potential
|
Isolate Code |
Form |
Elevation |
Edges |
Gram |
Catalase |
Oxidase |
IP |
|
SAT.2 |
Circular |
Convex |
Entire |
+ |
+ |
+ |
4,2 |
|
SAT.7 |
Filamentous |
Umbonate |
Filalmentous |
+ |
- |
- |
6,0 |
|
SAT.11 |
Circular |
Umbonate |
Entire |
+ |
+ |
- |
4,2 |
|
SAT.12 |
Circular |
Pulvinate |
Entire |
+ |
- |
- |
5,5 |
|
SET.10 |
Irregular |
Flat |
Undulate |
+ |
+ |
+ |
4,2 |
Description: SAT.2: Sediments block A center isolate 2, SAT.7: Sediments block A center isolate 7, SAT.11: Sediments block A middle isolate 11, SAT.12: Sediments block A middle isolate 12, SET.10: Sediments block E middle isolate 10; IP: Proteolytic Index.
Among the 97 bacteria isolates capable of producing proteolytic enzymes, 5 exhibited the highest activity index, namely SAT.2, SAT.7, SAT.12, SAT.11, and SET.10. Specifically, isolates SAT.2 and SET.10 showed positive Gram staining as well as catalase and oxidase activity, while SAT.7 and SAT.12 exhibited positive Gram staining, negative catalase, and oxidase activity. Isolate SAT.11 was positive Gram, with positive catalase, and negative oxidase activity. The five bacteria isolates showed great potential as degraders of organic matter contained in rice-fish farming system sediments. However, their precise identity was unknown, indicating the need for further biochemical and molecular characterization.
Based on the study results, there was a difference in the abundance of bacteria between Blocks A and E, where the abundance of bacteria in Block E was higher, namely around 2.0 - 9.2 high organic material content. Proteolytic bacteria form different proportions in the two blocks despite their relatively high bacterial abundances (Table 4). The highest proportion of proteolytic bacteria was found at the middle point of the two blocks, namely in Block A (76%) and Block E (72%). This difference in proportion is related to the accumulation of organic material with high protein content which is carried by water currents and settles in the middle of the pond. The high proportion of proteolytic bacteria found in sediment indicates a faster degradation process of organic material in the pond. This research also obtained five bacterial isolates that had the highest proteolytic activity index (>4), namely SAT.2, SAT.7, SAT.12, SAT.11, and SET.10. The high activity of the protease enzyme produced by the five isolates obtained shows its high ability to degrade proteins found in organic materials. These five isolates have the potential to convert organic waste and fish waste into probiotic agents that can break down organic compounds in the environment.
This research was funded by the Directorate General of Higher Education, Research and Technology, Ministry of Education, Culture, Research and Technology of the Republic of Indonesia through Applied Research - Downstream Path.
[1] Rahmadi, P.Z., Zuber, A., Wijaya, M. (2019). Case analysis of the role of self-help extension workers in the development of rice-fish farming systems in nogotirto village. Journal of Actual Progressive Sociological Research, 1(1): 94-106. https://doi.org/10.29303/resiprokal.v1i1.8
[2] Ibrahim, L.A., Shaghaleh, H., Abu-Hashim, M., Elsadek, E.A., Hamoud, Y.A. (2023). Exploring the integration of rice and aquatic species: Insights from global and national experiences. Water, 15(2750): 1-23. https://doi.org/10.3390/w15152750
[3] Li, F., Sun, Z., Qi, H., Zhou, X., Xu, C., Wu, D., Fang, F., Feng, J., Zhang, N. (2019). Effects of rice-fish co-culture on oxygen consumption in intensive aquaculture pond. Rice Science, 26(1): 50-59. https://doi.org/10.1016/j.rsci.2018.12.004
[4] Nursandi, J., Verdian, A.H. (2022). Minapadi Media Design for Fish and Rice Cultivation Systems in Dry Land. In IOP Conference Series: Earth and Environmental Science, 1012(1): 1-9. https://doi.org/10.1088/1755-1315/1012/1/012076
[5] Herlambang, A., Murwantoko, M., Istiqomah, I. (2021). Dynamic change in bacterial communities in the integrated rice-fish farming system in Sleman, Yogyakarta, Indonesia. Aquaculture Research, 52(11): 5566-5578. https://doi.org/10.1111/are.15432
[6] Satyantini, W.H., Salsabila, M., Nindarwi, D.D., Sahidu, A.M., Mukti, A.T. (2020). Provision of bacteria from shrimp pond sediment towards N/P ratio, plankton abundance, and total bacteria in the culture media of white shrimp (Litopenaeus vannamei). IOP Conference Series: Earth and Environmental Science, 441(1). https://doi.org/012139.https://doi.org/10.1088/1755-1315/441/1/012139
[7] Arunrat, N., Sansupa, C., Kongsurakan, P., Sereenonchai, S., Hatano, R. (2022). Soil microbial diversity and community composition in rice-fish co-culture and rice monoculture farming system. Biology, 11(8): 1-17. https://doi.org/10.3390/biology11081242
[8] MacKay, K.T. (1995). Rice-fish culture in China. In International Development Research Centre, Ottawa, Canada.
[9] Mufaidah, Z., Supriharyono, S., Muskananfola, M.R. (2016). Relationship between organic material content and total bacteria in the sediment of the wiso river estuary, Jepara. Management of Aquatic Resources Journal, 5(4): 265-274. https://doi.org/10.14710/marj.v5i4.14419
[10] Beristain, B.T. (2005). Organic Matter Decomposition in Simulated Aquaculture Ponds. Wageningen Institute of Animal Sciences, Wageningen University, Netherlands. https://edepot.wur.nl/121646.
[11] Li, Y., Li, L., Yu, Y., Hu, Q., Li, X. (2021). Impact of dietary protein content on soil bacterial and fungal communities in a rice-crab co-culture system. Frontiers in Microbiology, 12: 1-9. https://doi.org/10.3389/fmicb.2021.696427
[12] Qudus, A.F., Abiodun, L.R. (2023). Screening and isolation of protease-producing bacteria from wastewater samples in Obafemi Awolowo University (OAU) Campus, Ile-Ife, Nigeria. bioRxiv, 2023(7): 1-23. https://doi.org/10.1101/2023.07.09.548249
[13] Wettstadt, S., Lamas, M.A. (2020). Role of regulated proteolysis in the communication of bacteria with the environment. Frontiers in Molecular Biosciences, 7(10): 1-13. https://doi.org/10.3389/fmolb.2020.586497
[14] Artha, O.A., Sudarno., Pramono, H., Sari, L.A. (2019). Identification of extracellular enzyme-producing bacteria (proteolytic, cellulolytic, and amylolytic) in the sediment of extensive ponds in Tanggulrejo, Gresik. IOP Conference Series: Earth and Environmental Science. 236(1): 012003. https://doi.org/10.1088/1755-1315/236/1/012003.
[15] Balume, I., Agumas, B., Musyoki, M., Marhan, S., Cadisch, G., Rasche, F. (2022). Potential proteolytic enzyme activities modulate archaeal and bacterial nitrifier abundance in soils differing in acidity and organic residue treatment. Applied Soil Ecology, 169(2021): 1-8. https://doi.org/10.1016/j.apsoil.2021.104188
[16] Rizaldi, R., Setyantini, W.H., Sudarno, S. (2018). Isolation and Characterization of Proteolytic Bacteria Associated with Seagrass (Enhalus acoroides) at Bama Beach, Baluran National Park, Situbondo, East Java. Jurnal Ilmiah Perikanan dan Kelautan, 10(1): 8-14. https://doi.org/10.20473/jipk.v10i1.8314
[17] Solanki, P., Putatunda, C., Kumar, A., Bhatia, R., Walia, A. (2021). Microbial proteases: Ubiquitous enzymes with innumerable uses. 3 Biotech, 11(10): 428. https://doi.org/10.1007%2Fs13205-021-02928-z
[18] Su, H., Xiao, Z., Yu, K., Huang, Q., Wang, G., Wang, Y., Liang, J., Huang, W., Huang, X., Wei, F., Chen, B. (2020). Diversity of cultivable protease-producing bacteria and their extracellular proteases associated to scleractinian corals. PeerJ, 3: 1-17. https://doi.org/10.7717/peerj.9055
[19] Utomo, P.B., Sudarno., Rahardja, B.S. (2019). Identification of proteolytic bacterial isolates in sediment ecosystem of gunung anyar mangrove forest, Surabaya. In IOP Conference Series: Earth and Environmental Science, 236(1): 012060. https://doi.org/10.1088/1755-1315/236/1/012060
[20] Hastuti, U.S., Nugraheni, F.S.A., Asna, P.M.A. (2017). Identification and determination of protein hydrolysis index in proteolytic bacteria from mangrove soil in Margomulyo, Balikpapan. Proceeding Biology Education Conference. 14(1): 265-270. https://jurnal.uns.ac.id/prosbi/article/view/17788.
[21] Kurniawati, A. (2015). Isolation, selection, and identification of extracellular enzyme-producing bacteria from the digestive tract and sediment of intensive vanname shrimp (Litopenaeus vannamei) cultivation ponds. Thesis, Airlangga University, Surabaya.
[22] Prayogo., Rahardja, B.S., Asshanti, A.N., Dewi, N.N., Santanumurti, M.B. (2018). Exploration of indigenous bacteria in an intensive aquaculture system of African catfish (Clarias sp.) in Banyuwangi, Indonesia. IOP Conference Series: Earth and Environmental Science, 137(1): 012085. https://doi.org/10.1088/1755-1315/137/1/012085
[23] Amin, M., Yusuf, M.F., Suarsini, E., Anugrah, F.A., Alita, E.W., Rachman, I., Matsumoto, T. (2019). Identification of indigene bacteria from waste water of Regional Public Hospitals in Pacitan. IOP Conference Series: Earth and Environmental Science, 230(2019): 1-10. https://doi.org/10.1088/1755-1315/230/1/012089
[24] Prihanto, A.A., Aninta, K.D., Trisnaningrum, S. (2021). Protease production from Bacillus sp. isolated from gastrointestinal tract of Catfish (Clarias sp.) with different medium. Journal of Aquaculture and Fish Health, 10(2): 186-191. https://doi.org/10.20473/jafh.v10i2.17606
[25] Mulyasari, M., Sunarno, M.T.D., Suryaningrum, L.H. (2018). Isolation, characterization, and identification of proteolytic bacteria to improve protein digestibility of fish feed ingredients. Indonesian Aquaculture Journal, 13(2): 83-93. https://doi.org/10.15578/iaj.13.2.2018.83-93
[26] Nurhafid, M., Syakuri, H., Oedjijono, O., Listiowati, E., Ekasanti, A., Nugrayani, D., Pramono, H. (2021). Isolation and molecular identification of proteolytic bacteria from the digestive tract of tilapia (Oreochromis niloticus) Cultivated in Banyumas Regency. Jurnal Perikanan Universitas Gadjah Mada, 23(2): 95. https://doi.org/10.22146/jfs.64072
[27] Mirizadeh, S., Yaghmaei, S., Nejad, Z.G. (2014). Biodegradation of cyanide by a new isolated strain under alkaline conditions and optimization by response surface methodology (RSM). Journal of Environmental Health Science and Engineering, 12(1): 1-9. https://doi.org/10.1186/2052-336X-12-85
[28] Madigan, M.T., Martinko, J.M. (2006). Brock Biology of Microorganisms (Eleventh Edition). Pearson; Prentice Hall.
[29] Holt, J.G., Krieg, N.R., Sneath, P.H.A., Staley, J.T., WIlliams, S.T. (1994). Bergey’s Manual Of Determinative Bacteriologi (Ninth Edition). Lippincott Williams & Wilkins.
[30] Khiftiyah, A.M., Hanifah, N.N., Bachruddin, M., Sholichah, M.A., Istiqomah, S., Dewi, S.R., Rahayu, T., Prasetya, I.A.W., Marjayandari, L., Aini, N., Tsana, I., Triwahyuni, D., Fatimah., Ni’matuzahroh, N.M. (2018). Isolation and screening of potential proteolytic and amylolytic microbes from Wonorejo mangrove forest soil, Surabaya, Indonesia. In AIP Conference Proceedings, 2019(1): 040014-1-040014-5. https://doi.org/10.1063/1.5061884
[31] Pramono, H., Noer, T.I., Firdaus, M.R.A., Sudarno., Laksmi, S., Mubarak, A.S. (2019). Bacterial endophytes from mangrove leaves with antibacterial and enzymatic activities. Malaysian Journal of Microbiology, 15(7): 543-553. https://doi.org/10.21161/mjm.190352
[32] Firliani, W., Agustien, A., Fuji, D., Febria, A., Mikrobiologi, L., Biologi, J. (2013). Characterization of thermophilic bacteria in producing neutral protease enzymes. Jurnal Biologi Universitas Andalas, 4(1): 9-14. https://doi.org/10.25077/jbioua.4.1.%25p.2015
[33] Sinatryani, D. (2014). Abundance of cellulolytic bacteria in the Gunung Anyar River Estuary, Surabaya and Bancaran Bangkalan. Thesis, Airlangga University, Surabaya.
[34] Sudewi, S., Ala, A., Baharuddin., farid, M. (2020). The isolation, characterization endophytic bacteria from roots of local rice plant Kamba in, Central Sulawesi, Indonesia. Biodiversitas Journal of Biological Diversity, 21(4): 1614-1624. https://doi.org/10.13057/biodiv/d210442
[35] Khatoon, H., Chavan, D.D., Anokhe, A., Kalia, V. (2022). Catalase test: A biochemical protocol for bacterial identification. AgriCos e-Newsletter, 3(1): 53-55. http://www.agricosemagazine.com.
[36] Hadi, S.N., Dewi, P.S., Kartini. (2019). Identification of the ultisol land indigenus bacteria from Banyumas Regency based on the characteristics of morphology, physiology and biochemistry. In IOP Conference Series: Earth and Environmental Science, 250(1): 1-9. https://doi.org/10.1088/1755-1315/250/1/012095
[37] Kasprijo, Riady, R.M., Santosa, M., Fitriadi, R., Wijaya, R. (2022). Isolation of proteolytic bacteria from pond sediments for the maintenance of an integrated system fish and rice. International Journal of Scientific Engineering and Applied Science (IJSEAS), 8(7): 62-71. www.ijseas.com.
[38] Kondo, R., Shigematsu, K., Kawahara, N., Okamura, T., Yoon, Y.H., Sakami, T., Yokoyama, H., Koizumi, Y. (2012). Abundance of sulphate-reducing bacteria in fish farm sediments along the coast of Japan and South Korea. Fisheries Science, 78(1): 123-131. https://doi.org/10.1007/s12562-011-0439-3
[39] Kilawati, Y., Maimunah, Y., Amrillah, A.M., Kartikasari, D.P., Bhawiyuga, A. (2022). Characterization of water quality using IoT (Internet of Things), plankton and expression of virus-like particles in vannamei shrimp ponds of different constructions. Aquaculture, Aquarium, Conservation & Legislation, 15(2): 912-926. http://www.bioflux.com.ro/aacl.
[40] Guan, W., Yin, M., He, T., Xie, S. (2015). Influence of substrate type on microbial community structure in vertical-flow constructed wetland treating polluted river water. Environmental Science and Pollution Research, 22: 16202-16209. https://doi.org/10.1007/s11356-015-5160-9
[41] Irene, D.S., Dirgayusa, I.G.N.P., Puspitha, N.L.P.R. (2020). Identification of bacteria that have the potential to degrade hydrocarbons from mangrove substrates with sandy, muddy and clay textures. Journal of Marine and Aquatic Sciences, 6(2): 175-184. https://doi.org/10.24843/jmas.2020.v06.i02.p4
[42] Sousa, A.M., Machado, I., Nicolau, A., Pereira, M.O. (2013). Improvements on colony morphology identification towards bacterial profiling. Journal of Microbiological Methods, 95(3): 327-335. https://doi.org/10.1016/j.mimet.2013.09.020
[43] Peng, C., Li, P. (2023). Abiotic and biotic effects on microbial diversity of small water bodies in and around towns. Sustainability, 15(10): 8151. https://doi.org/10.3390/su15108151
[44] Wang, C., Michalet, R., Liu, Z., Jiang, X., Wang, X., Zhang, G., An, L., Chen, S., Xiao, S. (2020). Disentangling large-and small-scale abiotic and biotic factors shaping soil microbial communities in an alpine cushion plant system. Frontiers in Microbiology, 11(925): 1-17. https://doi.org/10.3389/fmicb.2020.00925
[45] Zuraidah, Wahyuni, D., Astuty, E. (2020). Morphological characteristics and activity test of thermophilic bacteria from the Ie Seuum (Hot springs) Tourist Area. Journal of Natural and Environmental Sciences, 11(2): 40-47. https://doi.org/10.20956/jal.v11i2.10200
[46] Suciati, P., Tjahjaningsih, W., Masithah, E.D., Pramono, H. (2016). Activity enzymatic of isolate lactic acid bacteria from the digestive tract of mud crab (Scylla Spp.) as a candidate probiotics. Jurnal Ilmiah Perikanan dan Kelautan, 8(2): 94-108. https://doi.org/10.20473/jipk.v8i2.11182
[47] Amin, M. (2018). Marine protease-producing bacterium and its potential use as an abalone probiont. Aquaculture Reports, 12: 30-35. https://doi.org/10.1016/j.aqrep.2018.09.004
[48] Mamangkey, J., Suryanto, D. (2021). Proteolytic activity of local bacteria isolated from crocodile raries. Jurnal Pro-Life, 8(2): 112-121. https://doi.org/10.33541/jpvol6Iss2pp102
[49] Fitriadi, R., Setyawan, A.C., Palupi, M., Nurhafid, M., Kusuma, R.O. (2023). Isolation and molecular identification of proteolytic bacteria from vaname shrimp (Lithopenaeus Vannamei) ponds as probiotic agents. Iraqi Journal of Veterinary Sciences, 37(1): 161-170. https://doi.org/10.33899/ijvs.2022.133468.2251
[50] Asril, M., Leksikowati, S.S. (2019). Isolation and selection of proteolytic bacteria from tofu liquid waste as a basis for determining agents for making biofertilizers. Elkawnie, 5(2): 86-99. https://doi.org/10.22373/ekw.v5i2.4356
[51] Istikhomah. (2021). Screening for proteolytic bacteria in sediment and pond water of vanname shrimp (Litopenaeus vannamei) in Bunton Village, Cilacap Regency. Thesis, General Soedirman University, Purwokerto.
[52] Paskandani, R., Ustadi, Husni, A. (2014). Isolation and utilization of proteolytic bacteria to improve the quality of milkfish presto processing wastewater. Journal of Human and Environment, 21(3): 310-316. https://doi.org/10.22146/jml.18558
[53] Admassie, M., Woldehawariat, Y., Alemu, T. (2022). In vitro evaluation of extracellular enzyme activity and its biocontrol efficacy of bacterial isolates from pepper plants for the management of phytophthora capsici. BioMed Research International, 2022: 1-13. https://doi.org/10.1155/2022/6778352
[54] Ginting, E.L. (2020). Screening and characterization of thermophilic proteolytic bacteria from moinit beach hot springs, North Sulawesi. PLATAX Scientific Journal, 8(1): 118-125. https://doi.org/10.35800/jip.8.1.2020.28628
[55] Fitriyanto, N.A., Hadi, S., Imam Bahtiyar, M., Adi Prasetyo, R., Erwanto, Y. (2020). Characterization and growth profile of proteolytic strain PK-4 isolated from local slaughterhouse wastewater. BIO Web of Conferences. 28: 1-4. https://doi.org/10.1051/bioconf/20202803001
[56] Bhandari, S., Poudel, D.K., Marahatha, R., Dawadi, S., Khadayat, K., Phuyal, S., Shrestha, S., Gaire, S., Basnet, K., Khadka, U., Parajuli, N. (2021). Microbial enzymes used in bioremediation. Journal of Chemistry, 2021: 1-17. https://doi.org/10.1155/2021/8849512
[57] Maron, P.A., Sarr, A., Kaisermann, A., Lévêque, J., Mathieu, O., Guigue, J., Karimi, B., Bernard, L., Dequiedt, S., Terrat, S., Chabbi, A., Ranjard, L. (2018). High microbial diversity promotes soil ecosystem functioning Pierre-Alain. Applied and Environmental Microbiology, 84(9): 1-13. https://doi.org/10.1128/AEM.02738-17b
[58] Wei, Y., Bu, J., Long, H., Zhang, X., Cai, X., Huang, A., Ren, W., Xie, Z. (2021). Community structure of protease-producing bacteria cultivated from aquaculture systems: Potential impact of a tropical environment. Frontiers in Microbiology, 12(2): 1-11. https://doi.org/10.3389/fmicb.2021.638129
[59] Ampou, E.E., Triyulianti, I., Nugroho, S.C. (2015). Associated bacteria on scleractinia corals in relation to the la-nina phenomenon on bunaken island. National Marine Journal, 10(2): 55. https://doi.org/10.15578/jkn.v10i2.6157
[60] Enggrob, K.L., Larsen, T., Peixoto, L., Rasmussen, J. (2020). Gram-positive bacteria control the rapid anabolism of protein-sized soil organic nitrogen compounds questioning the present paradigm. Scientific Reports, 10(1): 1-9. https://doi.org/10.1038/s41598-020-72696-y
[61] Susmitha, S., Gowri, R.S., Meenambigai, P., Ramitha, R., Vijayaraghavan, R. (2015). Physiochemical properties of purified Catalase enzyme from Azolla. International Journal of Current Microbiology and Applied Sciences, 4(8): 836-844. https://doi.org/10.20546/ijcmas.2016.502.095
[62] Erttmann, S.F., Gekara, N.O. (2019). Hydrogen peroxide release by bacteria suppresses inflammasome-dependent innate immunity. Nature Communications, 10(3493): 1-13. https://doi.org/10.1038/s41467-019-11169-x
[63] Andrés, C.M.C., Pérez de la Lastra, J.M., Juan, C.A., Plou, F.J., Pérez-Lebeña, E. (2022). Chemistry of hydrogen peroxide formation and elimination in mammalian cells, and its role in various pathologies. Stresses, 2(3): 256-274. https://doi.org/10.3390/stresses2030019
[64] González, H.S., Ghneim-Herrera, T. (2021). Heavy metals in soils and the remediation potential of bacteria associated with the plant microbiome. Frontiers in Environmental Science, 9(604216): 1-17. https://doi.org/10.3389/fenvs.2021.604216
[65] Hederstedt, L. (2022). Diversity of cytochrome c oxidase assembly proteins in bacteria. Microorganisms, 10(926): 1-17. https://doi.org/10.3390/microorganisms10050926
[66] Borisov, V., Nastasi, M.R., Forte, E. (2023). Cytochrome bd as Antioxidant Redox Enzyme. Molecular Biology, 1-8. https://doi.org/10.1134/S002689332306003
[67] Wang, X., Clément, R., Roger, M., Bauzan, M., Mazurenko, I., Poulpiquet, A.D., Ilbert, M., Lojou, E. (2019). Bacterial respiratory chain diversity reveals a cytochrome c oxidase reducing O2 at low overpotentials. Journal of the American Chemical Society, 141(28): 11093-11102. https://doi.org/10.1021/jacs.9b03268
[68] Khademian, M., Imlay, J.A. (2017). Escherichia coli cytochrome c peroxidase is a respiratory oxidase that enables the use of hydrogen peroxide as a terminal electron acceptor. In Proceedings of the National Academy of Sciences, 114(33): E6922-E6931. https://doi.org/10.1073%2Fpnas.1701587114
[69] Chavan, D.D., Anokhe, A., Kalia, V. (2022). Oxidase test: A biochemical methods in bacterial identification dharmappa. AgriCos e-Newsletter, 3(1): 31-33. http://www.agricosemagazine.com.