Assem Assylbek*![]() | Yelena Rakhimova
| Yelena Rakhimova![]() | Gulnaz Sypabekkyzy
| Gulnaz Sypabekkyzy![]() | Lyzzat Kyzmetova
| Lyzzat Kyzmetova![]() | Balnur Dzhunuskanova
| Balnur Dzhunuskanova![]()
©2023 IIETA. This article is published by IIETA and is licensed under the CC BY 4.0 license (http://creativecommons.org/licenses/by/4.0/).
OPEN ACCESS
This study aimed to assess the antagonistic activity of Trichoderma strains, isolated from the soils of Southeast Kazakhstan, against phytopathogenic fungi and their potential effects on the growth of two pea plant varieties, Nikitka and Zhigalova 112. A double culture method and paper disk inoculation were employed to evaluate the antagonistic activity of the Trichoderma strains. Comparisons of colony radius sizes indicated that T. longibrachiatum exhibited significant overlap with all test subjects, while the colony radii of T. harzianum and T. viridescens, although smaller than that of T. longibrachiatum, surpassed those of all test objects. The zones of growth suppression for phytopathogenic fungi ranged from 2.5±0.14 mm to 17.7±0.23 mm. Notably, the most pronounced suppression was observed in T. koningiopsis and T. viridescens strains. The Trichoderma strains demonstrated varied impacts on the growth of two pea plant varieties. Generally, an enhancement in root system growth was observed. This observation implies that the isolated Trichoderma strains, exhibiting high antagonistic activity against phytopathogenic fungi and stimulating pea root system growth, hold potential utility in sustainable agricultural practices.
Trichoderma, antagonistic activity, Kungei Alatau, phytopathogen, rhizosphere, soil fungus, strain
Agricultural productivity is frequently compromised by the detrimental effects of phytopathogenic fungi, which present significant challenges to crop health and yield. Traditionally, the management of these pathogens has relied on the use of chemical fungicides, an approach being increasingly debated due to its environmental implications, health risks, and the emergence of fungicide-resistant fungal strains. An alternative solution is suggested by the biocontrol properties of Trichoderma strains, whose antagonistic activity against soil-borne phytopathogenic fungi is well-documented. However, their potential utility in the unique ecological context of Southeast Kazakhstan remains under investigated.
Trichoderma strains offer promise for sustainable agriculture, both in the mitigation of phytopathogenic fungi and the promotion of plant growth. Understanding these strains' antagonistic activities and their effects on crop growth could contribute to environmentally friendly and potentially more effective alternatives to chemical fungicides. This is particularly pertinent in regions like Southeast Kazakhstan, where distinctive soil characteristics and climatic conditions may significantly influence interactions between crops, phytopathogens, and biocontrol agents such as Trichoderma. By examining the Trichoderma strains isolated from Southeast Kazakhstan soils, we can not only enhance our understanding of their antagonistic activities against phytopathogenic fungi but also study their effect on crop growth – in this case, peas. This research could contribute to the development of region-specific biocontrol strategies and may set a precedent for similar studies in other regions with unique agricultural contexts. Ultimately, this contributes to the broader goals of improving crop health, increasing agricultural productivity, and promoting sustainable farming practices [1, 2].
Types of the genus Trichoderma are among the most studied mushrooms [3, 4], since they are the producers of enzymes, antibiotics, and toxins used in obtaining drugs for biological control of plant diseases, to stimulate their growth, to improve the phytosanitary condition of the soil and reduce the accumulation of infectious potential in it [5-7]. Individual strains of the genus Trichoderma are able to control various phytopathogens that cause the oppression and death of many agricultural plants; such as Alternaria alternata, Phytophthora spp., Sclerotium cepivorum, Macrophomina phaseolina, Fusarium oxysporum, Rhizoctonia solani, and Pythium sp. and others on a wide range of plants [8-14].
On the territory of the Russian Federation, studies of the genus Trichoderma were carried out in three areas: in the field of practical application, ecology, and geographical distribution. The geographical distribution of species of the genus Trichoderma in certain regions of Russia was studied, 22 species of fungi of this genus were described and a key for identification was proposed [15, 16]. The authors noted species with a wide distribution and species characteristic of local regions. It has been established that representatives of Trichoderma are found mainly in forest ecosystems. 73% of all Trichoderma isolates were recovered from forest soil, forest litter, and deadwood, while only 13% were found in meadow and steppe soils [17-19]. The remaining 14% were recovered from the soils of agrocenoses and anthropogenic habitats [20]. In the field of practical application, strains for the creation of new biological products have been proposed [21, 22].
Despite the noted practical and ecological significance of Trichoderma strains [2, 23] and their global presence in soils across various climatic zones, comprehensive data on species diversity in Kazakhstan is notably lacking [24, 25]. Only recently have reports emerged on the presence of Trichoderma species in the soils of Central Kazakhstan, Alatau, and Kungey Alatau (Northern Tien Shan).
The current study was motivated by these gaps in our knowledge of Trichoderma strains in Southeast Kazakhstan and their antagonistic activity against prevalent soil phytopathogenic fungi. Moreover, the specific impact of these fungi on crop growth, especially peas, in the unique conditions of this region requires further investigation.
The aim of this study is to isolate Trichoderma strains from the soils of Southeast Kazakhstan and evaluate their antagonistic activity against prevalent phytopathogenic fungi. Additionally, the impact of these Trichoderma strains on the growth of peas, a common regional crop, will be examined. This research seeks to answer the following key questions: What Trichoderma strains can be isolated from Southeast Kazakhstan soils, and how antagonistic are they against the region's prevalent phytopathogenic fungi? How do these strains affect the growth of peas, specifically their aboveground and belowground development? Are there observable variations in the effectiveness of different Trichoderma strains, and could these strains serve as biocontrol agents for sustainable agricultural practices in the region?
The materials for the study were obtained during the implementation of the project "Obtaining domestic strains of fungi of the genus Trichoderma for soil improvement in agrobiocenoses" (IRN AR08052881).
Soil samples were taken on the territory of the Kungei Alatau ridge, belonging to the Northern Tien Shan Mountain system and located in the southeastern part of Kazakhstan. Sampling was carried out at the following points: gorge Taldy, along the Taldy River, floodplain deciduous forest, strain 307p, 307r, 1694 m above sea level, N43°013333.9ʺ, EO78°15ʹ29.6ʺ; the road from the gorge Kudurga to the gorge Kurmety, strain 324j, 1571 m above sea level, N43°024646.6ʺ, EO78°18ʹ34.5ʺ, gorge Kolsai, the road from the Saty village to the Lower Kolsai lake, deciduous forest, strain 330v, 1550 m above sea level, N43°023333.5ʺ, EO78°20ʹ38.5ʺ. Samples were taken in the soil layer of 5-20 cm of mountain forest soils, in the rhizosphere of wormwood (Artemisia sp.), mountain ash (Sorbus tianschanica Rupr.), spiraea (Spiraea hypericifolia L.), poplar (Populus talassica Kom.).
For the study, a total of 50 soil samples were collected from the selected sampling points, with each point being sampled 10 times, thus ensuring robustness and diversity in the data collected. The selection of these sites was based on their varied altitudes, diverse vegetation, and distinct soil types, factors that potentially influence the variety of Trichoderma strains present in the soil. The choice of the Northern Tien Shan Mountain system, in particular, was due to its being a less-studied region, hence the potential to discover novel or endemic strains of Trichoderma fungi. As for the plant species chosen for the rhizosphere sampling, wormwood (Artemisia sp.), mountain ash (Sorbus tianschanica Rupr.), spiraea (Spiraea hypericifolia L.), and poplar (Populus talassica Kom.) were selected due to their abundance and ecological importance in the region. These plants are known to interact symbiotically with various fungi, hence the interest in studying the fungi in their rhizosphere. Also, the rhizosphere of these plants was thought to offer a favorable environment for Trichoderma, hence enhancing the potential to isolate a diverse range of strains. These strategic decisions guided our sampling process, aiming to maximize the study's yield in terms of both quantity and quality of Trichoderma strains isolated.
For the isolation and further identification of soil fungi common methods were used [26]. Potato-sucrose agar was used to isolate fungi of the genus Trichoderma [27-29]. All samples were inoculated at a temperature of +25-27℃ [30, 31].
Three to seven fungal strains each day were employed to identify mushroom samples by molecular genetics. At -20℃, the mycelium was frozen. then pounded with a pestle until powdery in a 1.5 ml Eppendorff test tube. Following the manufacturer's instructions, DNA was extracted from the resultant mass using Norgen Biotek Corp's (Ontario, Canada) Plant/Fungi DNA Isolation Kit. DNA was extracted from the acquired masses in accordance with the manufacturer's instructions using the Plant/Fungi DNA Isolation Kit from Norgen Biotek Corp (Ontario, Canada). The dsDNA HS scale was used to calculate the DNA concentration in the samples using a Qubit TM dsDNA HS Assay Kit fluorimeter (Life Technologies, Oregon, USA). Primers for the ITS region of all fungi were utilized, specifically ITS1 (5-TCCGTAGTAGGTGAACCTGCGG-3) and (5-TCCTCCGCTTATTGATATGC-3) [32]. The reaction mixture for amplification contained 1.5 ml of DNA, 1.25 ml of forward primer (10 M), 1.25 ml of reverse primer (10 M), 12.5 ml of Q5® Hot Start High-Fidelity 2X Master Mix, and 8.5 water [33]. The polymerase chain reaction (PCR) mixture afterwards had a mass of 25 L [34].
The following amplification modes were used during PCR using an Eppendorf ProS amplifier (Hamburg, Germany): 94℃ for 30 sec, 55℃ for 1 min, 72℃ for 40 sec, for a total of 30 cycles, and 72℃ for 10 min. On a 1.2% agarose gel, the resulting amplification data were examined. Utilizing the CleanSweepTM PCR Purification reagent, PCR products were purified (Applide Biosystems, USA). According to the manufacturer's instructions (BigDye® Terminator v3.1 Cycle Sequencing Kit Protocol Applied Biosystems USA), the sequencing reaction was carried out using the BigDye Terminator v3.1 Cycle Sequencing Kit from Applied Biosystems, USA. Further fragment separation was performed using a 3500 automatic DNA analyzer (Applied Biosystems, USA) [32]. SeqA software was used to process the sequencing findings (Applied Biosystems, USA). Using the BLAST tool, the nucleotide sequences of the ITS region of fungi were compared with the GeneBank database [35].
Phylogenetic trees were constructed using the Neighbour-Joining (NJ) algorithm and aligned using the ClustalW algorithm. Building the phylogenetic trees using the Neighbour-Joining (NJ) method was an integral part of our analysis. For this process, we used the Tamura-Nei model as our substitution model, which is commonly used for nucleotide substitution in bioinformatics studies due to its ability to accommodate unequal base frequencies and different transition and transversion rates. The model selection was based on the lowest Bayesian Information Criterion (BIC) scores, an indication of the optimal balance between the goodness of fit of the model and its complexity. To evaluate the statistical support for each branch, we conducted 1000 bootstrap replications, which provided us with a percentile estimate of the reliability of each branch's positioning within the tree. Branches with bootstrap values above 70% were generally considered as statistically supported. For the construction of the phylogenetic trees, we used an outgroup to root the tree, which was a well-identified sequence of a closely related species to Trichoderma, but distinctly separate from it. This helped in providing a clearer evolutionary relationship among the Trichoderma strains we studied.
Antagonistic properties of fungi of the genus Trichoderma in relation to phytopathogenic fungi Alternaria sp., Aspergillus niger, Fusarium sp. (strains 376j, 320, 323), Penicillium expansum, and Purpureocillium lilacinum, stored in the collection of the Institute of Botany and Phytointroduction, were determined by the method of double cultures at the optimal temperature for the growth of microorganisms 25-26℃ on potato-sucrose agar. The experiments were carried out in three variants:
1. Trichoderma fungus (strains: 307p, 307r, 324j, 330v) was sown with the pathogen simultaneously;
2. Introduction of Trichoderma (strains: 307p, 307r, 324j, 330v) to the pathogen after two days;
3. Introduction of Trichoderma (strains: 307p, 307r, 324j, 330v) to the pathogen after four days.
The size of fungal colonies was measured on 4-6 days by the linear method (in mm) from the back of the cup.
The inhibitory activity of Trichoderma fungi (strains: 307p, 307r, 324j, 330v) was determined by paper disks. A disk of filter paper moistened with an extract from the Trichoderma mycelium was applied to the test culture of phytopathogenic fungi evenly distributed over the surface of the solid nutrient medium in a Petri dish. After incubation, the growth suppression zone of the test culture was determined on the fifth day. The Trichoderma mycelium extract was prepared through a series of steps. Initially, the Trichoderma strains were cultivated in a nutrient-rich broth until an abundant growth of mycelium was observed, which took approximately 14 days. Once optimal growth was achieved, the mycelium was carefully harvested using a sterile loop and rinsed with sterile distilled water to remove any residual medium. The clean mycelium was then ground using a mortar and pestle in the presence of methanol, acting as a solvent to facilitate the extraction of bioactive compounds. After grinding, the mixture was filtered using Whatman filter paper to obtain a clear extract, which was then subjected to rotary evaporation at 40 degrees Celsius to remove methanol. This process resulted in a concentrated extract. For the inhibition test, a volume of 50 microliters of the extract was applied to each filter paper disk, providing a sufficient amount to moisten the disk without causing it to disintegrate.
Isolates of Trichoderma fungi (strains: 307p, 307r, 324j, 330v) and pea plants (varieties Nikitka, Zhigalova 112) were used as objects of research. Seeds that were not treated with spores were used in the control. Seeds were sown in a nutrient soil for seedlings Volshebnaya Gryadka (Buisk fertilisers) in mini cassettes for seedlings with a cell size of 55 × 55 × 40 (Agrocom 2015). On the 7th-8th day after germination, the length of stems and roots was measured.
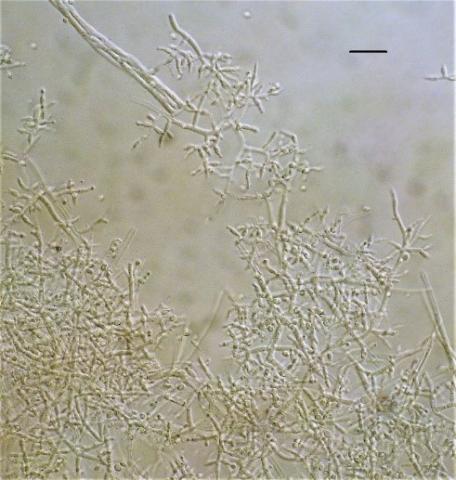
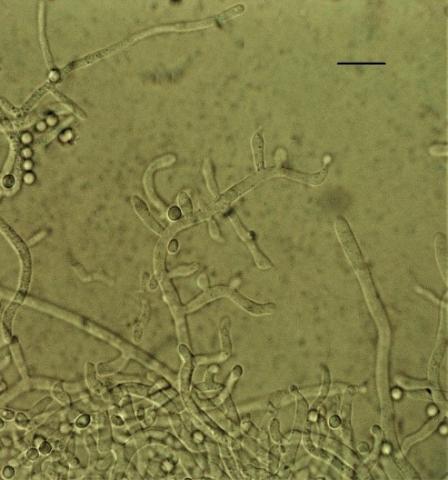
From soil samples of the rhizosphere of plants of the Kungei Alatau ridge, 4 species of the genus Trichoderma were identified by morphological and cultural characteristics and sequencing: T. viridescens (A.S. Horne & H.S. Will.) Jaklitsch & Samuels, T. koningiopsis Samuels, Carm. Suárez & H.C. Evans (Figure 1), T. harzianum Rifai, T. longibrachiatum Rifai (Figure 2). According to some researchers [15, 36, 37], the last three species are characteristic of the southern regions.
Figure 1. Conidiophores Trichoderma koningiopsis (scale 15 microns)
Figure 2. Conidiophores Trichoderma longibrachiatum (scale 10 microns)
When comparing the received nucleotide sequences of the ITS region of fungal DNA with the GeneBank, 99.62% similarity was revealed in Trichoderma koningiopsis isolated from the rhizosphere of Spiraea hypericifolia, and 100% similarity in the other three species of T. viridescens, T. harzianum, T. longibrachiatum (Table 1).
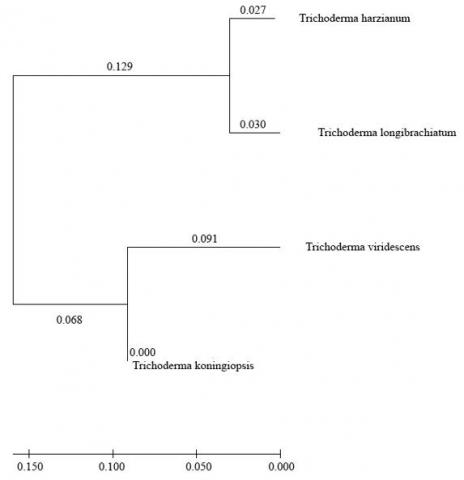
Four Trichoderma strains in a built phylogenetic tree with nucleotide sequences of ITS DNA region were divided into two distinct groups (Figure 3). Group 1 included isolates of Trichoderma harzianum and T. longibrachiatum; group 2 included isolates of T. koningiopsis and T. viridescens.
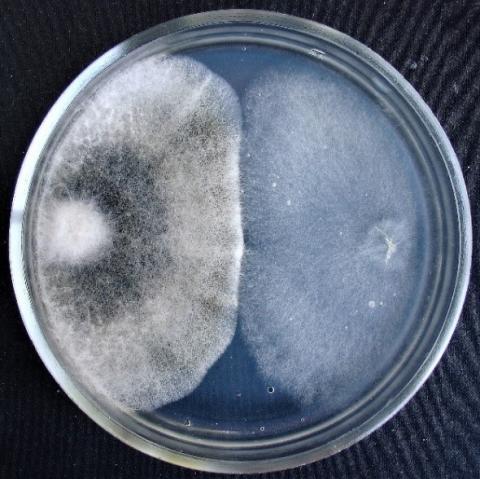
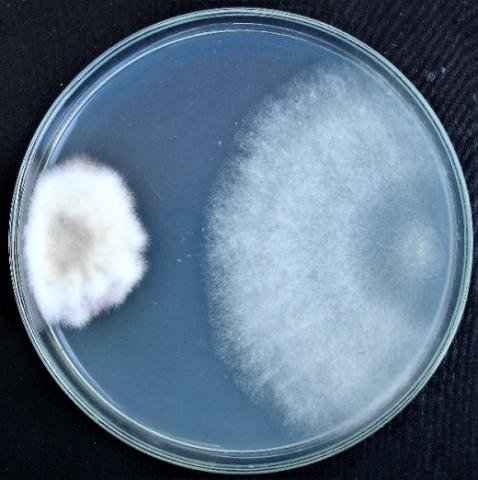
When determining the inhibitory activity of Trichoderma viridescens strain 307p by the method of double cultures, it was found that with simultaneous inoculation of Trichoderma and test objects on Petri dishes, the rapid growth of T. viridescens is observed in all variants (Table 2, Figures 4-7).
On the sixth day, the colony radius reaches the highest values in the variant with Purpureocillium lilacinum, Fusarium sp. (strain 376j) and Alternaria sp. (59.7±1.77 mm; 56.7±2.08 mm; 54.7±0.95 mm, respectively). In the case of the second variant of the experiment (T. viridescens inoculation after 2 days), the radius of the Trichoderma colonies overlaps the radius of the test objects in the case of Fusarium spp. (strains 376j, 320, 323) and Penicillium expansum. In the third variant of the experiment, only in one case does the radius of the Trichoderma colony overlap the radius of the test object: when using Penicillium expansum as a test object.
Table 1. Results of determination of representatives of the genus Trichoderma, BLAST, NCBI
|
Strains |
Similarity % |
Access Number GenBank |
Organism GenBank |
|
307p (from the rhizosphere of Artemisia sp.) |
100.00% |
MK804405.1:33-496 |
Trichoderma viridescens |
|
307r (from the rhizosphere Sorbus tianschanica Rupr.) |
100.00% |
MK738147.1:113-606 |
Trichoderma harzianum |
|
324j (from the rhizosphere of Spiraea hypericifolia L.) |
99.62% |
KJ191357.1:2-261 |
Trichoderma koningiopsis |
|
330v (from the rhizosphere of Populus talassica Kom.) |
100.00% |
MT520608.1:102-611 |
Trichoderma longibrachiatum |
Table 2. Determination of the inhibitory activity of the Trichoderma viridescens strain (307p) by the method of double cultures
|
No. |
Test Object |
Colony Radius in mm in Different Variants |
|||||
|
Simultaneous Inoculation |
Inoculation after 2 Days |
Inoculation after 4 Days |
|||||
|
On the 4th Day |
On the 6th Day |
On the 4th Day |
On the 6th Day |
On the 4th Day |
On the 6th Day |
||
|
1 |
T. viridescens |
33.9±1.60 |
54.7±0.95 |
18.3±0.69 |
37.1±0.54 |
- |
18.8±1.07 |
|
Alternaria sp. |
24.7±0.75 |
28.8±1.06 |
28.2±0.44 |
40.8±0.64 |
41.3±0.40 |
52.5±0.97 |
|
|
2 |
T. viridescens |
42.7±3.43 |
52.1±3.13 |
21.9±0.94 |
30.9±1.69 |
- |
13.4±1.14 |
|
Aspergillus niger |
34.4±2.13 |
37.0±4.26 |
54.6±0.59 |
62.4±1.96 |
65.4±1.02 |
70.3±0.55 |
|
|
3 |
T. viridescens |
43.9±2.43 |
56.7±2.08 |
18.5±1.10 |
38.3±0.53 |
- |
21.2±1.45 |
|
Fusarium sp. (strain 376 w) |
6.5±0.68 |
11.7±0.57 |
17.7±0.45 |
19.3±1.77 |
17.6±0.49 |
21.4±0.55 |
|
|
4 |
T. viridescens |
30.7±0.79 |
42.2±4.10 |
19.2±0.39 |
37.3±1.81 |
- |
19.1±0.36 |
|
Fusarium sp. (strain 320) |
21.9±2.52 |
29.5±1.08 |
24.6±0.70 |
28.5±1.09 |
29.2±1.61 |
35.8±0.34 |
|
|
5 |
T. viridescens |
17.4±1.34 |
45.1±3.90 |
24.6±1.28 |
38.2±0.49 |
- |
16.3±0.38 |
|
Fusarium sp. (strain 323) |
8.0±0.65 |
10.9±0.64 |
23.1±2.52 |
24.8±0.54 |
15.9±0.49 |
18.7±0.51 |
|
|
6 |
T. viridescens |
42.1±2.17 |
51.7±2.55 |
27.8±3.34 |
40.7±0.53 |
- |
18.4±1.37 |
|
Penicillium expansum |
5.4±0.38 |
9.1±0.43 |
8.5±1.56 |
11.7±1.30 |
8.8±1.19 |
15.9±0.46 |
|
|
7 |
T. viridescens |
30.9±3.70 |
59.7±1.77 |
20.1±0.94 |
23.1±2.57 |
- |
9.9±1.55 |
|
Purpureocillium lilacinum |
9.9±0.10 |
15.8±2.27 |
25.2±2.16 |
26.5±0.56 |
14.1±1.00 |
15.5±0.99 |
|
Figure 3. Phylogenetic relationships of four isolated Trichoderma species obtained by ITS-region DNA analysis
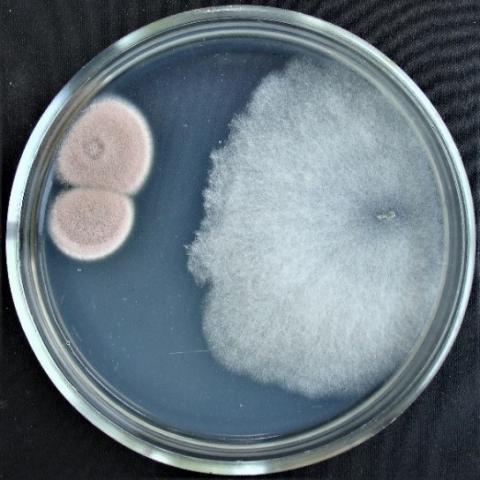
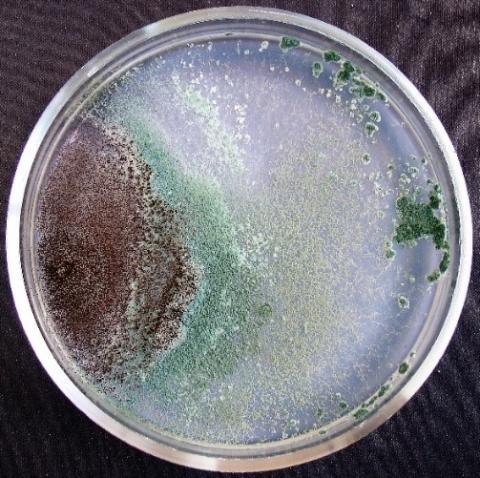
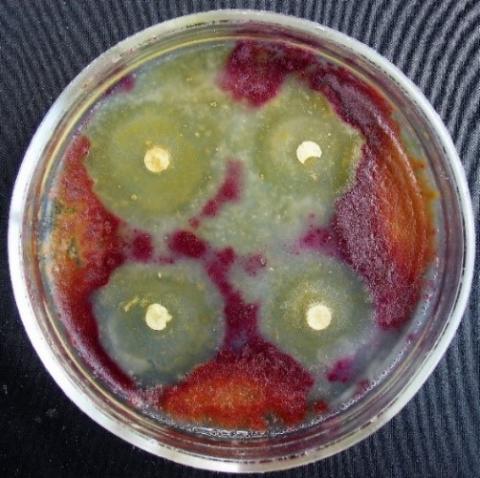
Figure 4. Double cultures of T. viridescens and Alternaria sp., simultaneous inoculation
Figure 5. Double cultures of T. viridescens and Aspergillus niger, simultaneous inoculation
Figure 6. Double cultures of T. viridescens and Fusarium sp. (strain 376j), simultaneous inoculation
Figure 7. Double cultures of T. viridescens and Fusarium sp. (strain 323), simultaneous inoculation
Table 3. Determination of the inhibitory activity of the Trichoderma koningiopsis strain (strain 324j) by the method of double cultures
|
No. |
Test Object |
Colony Radius in mm in Different Variants |
|||||
|
Simultaneous Inoculation |
Inoculation after 2 Days |
Inoculation after 4 Days |
|||||
|
On the 4th Day |
On the 6th Day |
On the 4th Day |
On the 6th Day |
On the 4th Day |
On the 6th Day |
||
|
1 |
T. koningiopsis |
38.9±0.94 |
47.9±1.17 |
21.6±1.10 |
41.9±0.41 |
- |
23.1±0.46 |
|
Alternaria sp. |
27.7±2.22 |
34.5±2.16 |
30.7±0.60 |
39.3±0.45 |
37.6±0.60 |
42.7±0.45 |
|
|
2 |
T. koningiopsis |
38.7±0.55 |
44.4±1.83 |
15.1±1.04 |
30.4±1.72 |
- |
15.8±1.29 |
|
Aspergillus niger |
38.5±0.51 |
47.4±2.19 |
46.0±0.55 |
61.2±1.91 |
64.7±0.98 |
68.0±0.73 |
|
|
3 |
T. koningiopsis |
38.3±0.66 |
49.6±0.51 |
49.1±1.35 |
67.9±1.24 |
- |
24.5±0.43 |
|
Fusarium sp. (strain 376 w) |
8.0±0.53 |
16.0±1.50 |
13.7±1.46 |
20.1±1.58 |
14.4±0.71 |
22.2±0.63 |
|
|
4 |
T. koningiopsis |
42.1±0.39 |
51.8±0.56 |
14.6±1.06 |
39.8±0.37 |
- |
23.4±0.29 |
|
Fusarium sp. (strain 320) |
29.2±1.69 |
33.4±1.63 |
14.7±0.25 |
24.2±0.34 |
25.4±1.03 |
33.4±0.57 |
|
|
5 |
T. koningiopsis |
37.1 ±0.48 |
46.5±0.45 |
23.0±0.63 |
43.1±0.64 |
- |
22.8±0.31 |
|
Fusarium sp. (strain 323) |
15.9±1.03 |
22.6±1.92 |
14.7±0.89 |
19.7±1.31 |
17.2±0.60 |
20.7±0.38 |
|
|
6 |
T. koningiopsis |
39.1±1.08 |
49.5±0.98 |
19.4±0.40 |
45.8±0.75 |
- |
23.3±0.25 |
|
Penicillium expansum |
9.9±0.50 |
10.2±0.55 |
6.3±0.32 |
11.7±0.99 |
7.4±0.53 |
9.0±0.91 |
|
|
7 |
T. koningiopsis |
38.5±1.06 |
46.3±2.22 |
23.9±0.54 |
37.7±1.54 |
- |
19.7±0.28 |
|
Purpureocillium lilacinum |
10.5±0.57 |
15.2±1.00 |
11.2±0.76 |
35.5±6.67 |
12.9±0.41 |
17.3±0.66 |
|
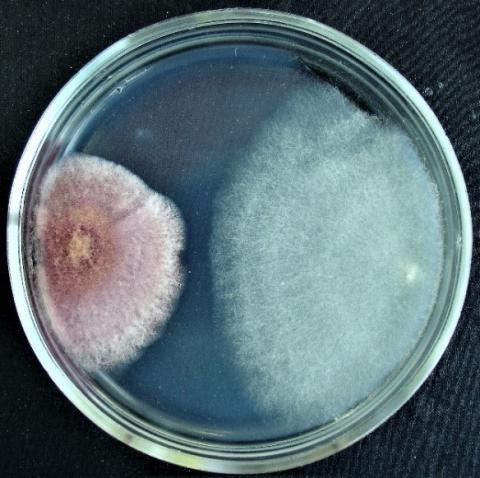
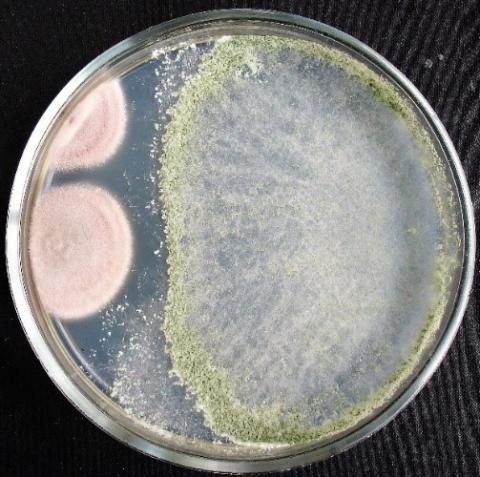
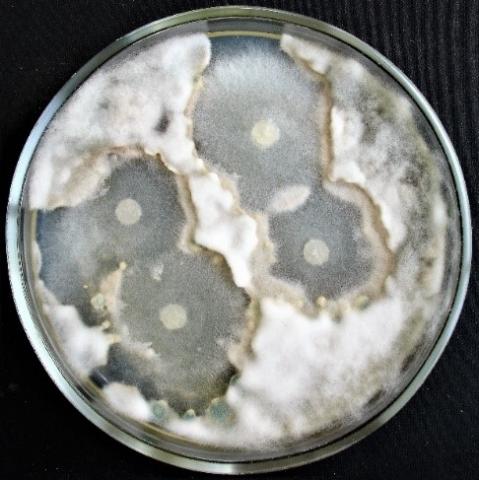
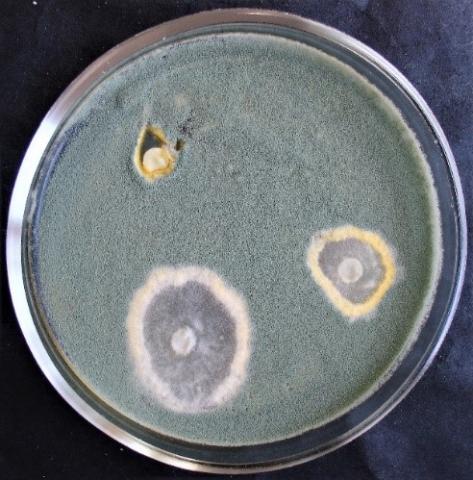
Figure 8. Double cultures of T. koningiopsis and Alternaria sp., simultaneous inoculation
Figure 9. Double cultures of T. koningiopsis and Fusarium sp. (strain 323), simultaneous inoculation
Figure 10. Double cultures of T. koningiopsis and Fusarium sp. (strain 320), simultaneous inoculation
Figure 11. Double cultures of T. koningiopsis and Purpureocillium lilacinum, simultaneous inoculation
With simultaneous inoculation of Trichoderma and test objects on Petri dishes to determine the inhibitory activity of Trichoderma koningiopsis strain 324 by the method of double cultures, it was found that the rapid growth of T. koningiopsis colonies is characteristic of almost all experimental variants (Table 3, Figures 8-11). The exception is the joint inoculation of T. koningiopsis and Aspergillus niger, in this case, the radius of aspergillus colonies on the sixth day is significantly larger (47.4±2.19 mm) than that of Trichoderma colonies. In other cases, on the sixth day, the colony radius reaches the highest values in the variant with Fusarium sp. (strain 320), Penicillium expansum and Fusarium sp. (strain 376j) (51.8±0.56 mm; 49.5±0.98 mm; 49.6±0.51 mm, respectively).
When the Trichoderma is introduced to the test object on the second day, the same pattern persists: the radius of the aspergillus colonies on the sixth day is significantly larger (61.2±1.91 mm) than that of the Trichoderma colonies, and the radius of the colonies is T. koningiopsis reaches the highest values in the variant with Fusarium sp. (strain 376j), Fusarium sp. (strain 320) and Penicillium expansum (67.9±1.24 mm; 43.1±0.64 mm; 45.8±0.75 mm, respectively).
When the Trichoderma is introduced to the test object on the fourth day, the radius of the Trichoderma colonies overlaps the radius of the test objects in the case of using Fusarium sp. (strain 376j), Fusarium sp. (strain 323), Penicillium expansum and Purpureocillium lilacinum as test objects.
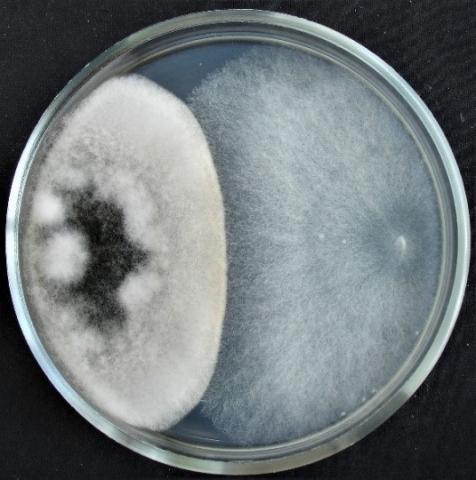
When determining the inhibitory activity of Trichoderma harzianum strain 307r by the method of double cultures, it was found that with simultaneous inoculation of Trichoderma and test objects on Petri dishes, rapid growth of T. harzianum colonies is observed in all variants of the experiment (Table 4, Figures 12 and 13). At the same time, on the sixth day, the radius of the Trichoderma colonies reaches the highest values in the variant with Penicillium expansum, Fusarium sp. (strain 320) and Aspergillus niger (66.4±1.49 mm; 55.4±5.28 mm; 54.8±2.91 mm, respectively).
In the variant of introduction of Trichoderma to the test object on the second day, the radius of the Trichoderma colonies overlaps the radius of all test objects, reaching the highest values with Penicillium expansum, Fusarium sp. (strain 376j) and Alternaria sp. (76.0±1.20 mm; 68.6±0.69 mm; 63.8±0.71 mm, respectively). When T. harzianum is introduced to the test object on the fourth day, the radius of the Trichoderma colonies overlaps the radius of the test objects in the case of using Fusarium sp. (strain 376j), Fusarium sp. (strain 323), Penicillium expansum and Purpureocillium lilacinum as test objects.
Table 4. Determination of the inhibitory activity of the Trichoderma harzianum strain (strain 307r) by the method of double cultures
|
No. |
Test Object |
Colony Radius in mm in Different Variants |
|||||
|
Simultaneous Inoculation |
Inoculation after 2 Days |
Inoculation after 4 Days |
|||||
|
On the 4th Day |
On the 6th Day |
On the 4th Day |
On the 6th Day |
On the 4th Day |
On the 6th Day |
||
|
1 |
T. harzianum |
29.0±1.41 |
41.2±1.90 |
59.6±1.12 |
63.8±0.71 |
- |
24.3±1.15 |
|
Alternaria sp. |
26.9±0.78 |
37.3±1.59 |
24.1±0.31 |
36.3±0.72 |
34.7±0.40 |
40.3±0.86 |
|
|
2 |
T. harzianum |
47.6±3.69 |
54.8±2.91 |
47.9±2.26 |
51.7±0.56 |
- |
11.4±0.89 |
|
Aspergillus niger |
16.8±0.58 |
27.7±2.13 |
46.2±1.68 |
48.0±1.64 |
67.4±0.47 |
71.1±1.51 |
|
|
3 |
T. harzianum |
21.1±0.54 |
46.0±2.33 |
63.5±3.40 |
68.6±0.69 |
- |
29.0±0.40 |
|
Fusarium sp. (strain 376 w) |
17.3±1.00 |
19.1±0.54 |
11.7±0.49 |
19.3±0.72 |
18.3±0.55 |
21.5±0.79 |
|
|
4 |
T. harzianum |
21.6±0.52 |
55.4±5.28 |
60.1±1.17 |
60.7±1.03 |
- |
31.2±0.60 |
|
Fusarium sp. (strain 320) |
9.8±1.20 |
33.9±2.09 |
18.7±0.57 |
22.0±0.42 |
28.5±3.10 |
36.8±0.51 |
|
|
5 |
T. harzianum |
30.4±0.41 |
45.9±3.39 |
24.6±0.83 |
25.8±0.54 |
- |
29.7±0.28 |
|
Fusarium sp. (strain 323) |
21.2±0.94 |
35.9±3.10 |
14.6±0.57 |
16.7±0.61 |
18.3±0.38 |
19.3±0.38 |
|
|
6 |
T. harzianum |
33.8±1.37 |
66.4±1.49 |
57.7±0.60 |
76.0±1.20 |
- |
25.9±1.96 |
|
Penicillium expansum |
11.3±0.58 |
32.5±2.43 |
7.3±0.35 |
9.5±0.25 |
5.5±0.5 |
6.6±0.67 |
|
|
7 |
T. harzianum |
37.8±2.39 |
43.2±2.42 |
55.8±0.96 |
58.7±1.01 |
- |
31.4±0.60 |
|
Purpureocillium lilacinum |
11.5±0.25 |
22.8±1.84 |
17.7±0.28 |
18.1±0.28 |
13.9±0.22 |
15.8±0.19 |
|
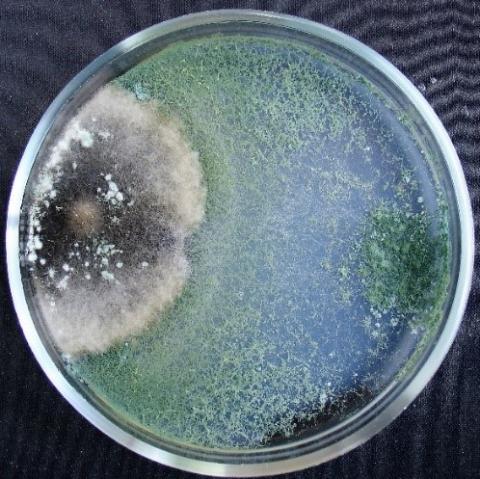
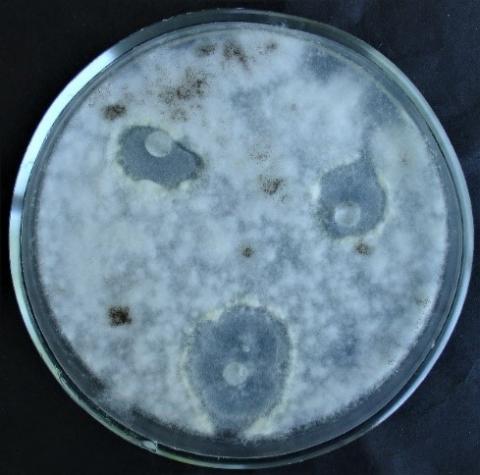
Figure 12. Double cultures of T. harzianum and Aspergillus niger, simultaneous inoculation
Figure 13. Double cultures of T. harzianum and Purpureocillium lilacinum
Table 5. Determination of the inhibitory activity of the Trichoderma longibrachiatum strain (strain 330v) by the method of double cultures
|
No. |
Test Object |
Colony Radius in mm in Different Variants |
|||||
|
Simultaneous Inoculation |
Inoculation after 2 Days |
Inoculation after 4 Days |
|||||
|
On the 4th Day |
On the 6th Day |
On the 4th Day |
On the 6th Day |
On the 4th Day |
On the 6th Day |
||
|
1 |
T. longibrachiatum |
24.0±2.22 |
48.9±5.56 |
54.2±0.48 |
59.6±2.87 |
- |
56.0±3.50 |
|
Alternaria sp. |
30.1±2.35 |
30.2±2.35 |
24.3±1.24 |
28.4±0.78 |
36.3±0.51 |
57.9±2.54 |
|
|
2 |
T. longibrachiatum |
34.1±4.47 |
37.7±2.60 |
64.0±3.02 |
68.3±0.38 |
- |
39.3±3.90 |
|
Aspergillus niger |
26.9±1.22 |
27.3±4.61 |
26.8±2.90 |
28.1±1.40 |
68.5±0.32 |
84.0±2.05 |
|
|
3 |
T. longibrachiatum |
8.5±1.00 |
59.6±2.54 |
68.9±2.21 |
69.4±1.82 |
- |
- |
|
Fusarium sp. (strain 376 w) |
5.9±0.64 |
17.3±1.10 |
12.7±0.57 |
13.5±0.47 |
- |
- |
|
|
4 |
T. longibrachiatum |
22.5±2.45 |
56.1±3.63 |
66.6±1.92 |
68.5±0.34 |
- |
53.4±2.33 |
|
Fusarium sp. (strain 320) |
21.5±1.24 |
30.1±2.42 |
16.8±0.80 |
25.1±0.75 |
30.6±0.29 |
65.9±2.77 |
|
|
5 |
T. longibrachiatum |
14.2±1.03 |
73.1±2.32 |
66.8±1.04 |
69.4±1.74 |
- |
58.5±1.83 |
|
Fusarium sp. (strain 323) |
10.8±0.85 |
20.7±0.98 |
12.8±0.58 |
18.3±0.83 |
18.1±0.79 |
30.6±2.48 |
|
|
6 |
T. longibrachiatum |
13.9±0.68 |
68.1±2.24 |
58.5±0.57 |
70.7±2.49 |
- |
53.9±1.17 |
|
Penicillium expansum |
8.7±1.04 |
23.6±1.16 |
7.2±0.62 |
8.0±0.18 |
6.1±0.25 |
17.1±1.04 |
|
|
7 |
T. longibrachiatum |
38.4±4.44 |
62.2±3.11 |
- |
- |
- |
7.0±0.77 |
|
Purpureocillium lilacinum |
10.0±0.73 |
12.8±0.49 |
- |
- |
10.0±0.72 |
17.5±1.63 |
|
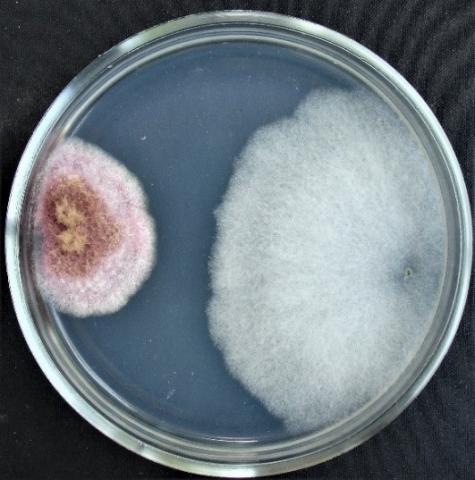
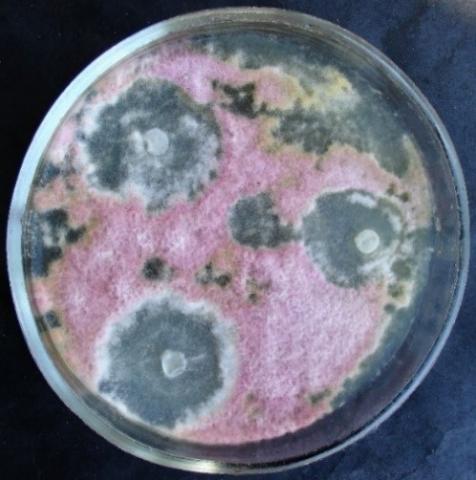
Figure 14. Double cultures of T. longibrachiatum and Alternaria sp., simultaneous inoculation
Figure 15. Double cultures of T. longibrachiatum and Aspergillus niger, simultaneous inoculation
Figure 16. Double cultures of T. longibrachiatum and Fusarium sp. (strain 323), simultaneous inoculation
Figure 17. Double cultures of T. longibrachiatum and Fusarium sp. (strain 320), simultaneous inoculation
Table 6. The zone of growth suppression of phytopathogenic fungi by extracts of representatives of the genus Trichoderma by the method of paper disks, on day 5, in mm
|
Test Object |
Growth Suppression Zone |
|||
|
T. viridescens |
T. koningiopsis |
T. harzianum |
T. longibrachiatum |
|
|
Alternaria sp. |
12.2±0.69 |
17.7±0.23 |
8.6±1.06 |
3.9±0.83 |
|
Aspergillus niger |
5.8±0.38 |
7.5±0.42 |
5.7±0.57 |
7.3±1.51 |
|
Fusarium sp. (strain 376j) |
16.4±0.48 |
12.5±1.24 |
9.3±1.00 |
- |
|
Fusarium sp. (strain 320) |
3.1±0.44 |
8.4±0.64 |
2.5±0.14 |
4.8±0.34 |
|
Fusarium sp. (strain 323) |
13.3±3.50 |
11.5±0.19 |
13.9±3.64 |
8.0±0.99 |
|
Penicillium expansum |
8.1±1.53 |
8.6±0.57 |
14.5±1.00 |
9.1±1.01 |
|
Purpureocillium lilacinum |
10.7±0.62 |
3.6±0.24 |
9.7±1.44 |
- |
With simultaneous inoculation of Trichoderma and test objects on Petri dishes to determine the inhibitory activity of Trichoderma longibrachiatum strain 330v by the method of double cultures, it was found that the rapid growth of T. longibrachiatum colonies is characteristic of all experimental variants (Table 5, Figures 14-17). At the same time, on the sixth day, the radius of the Trichoderma colonies reaches the highest values in the variant with Fusarium sp. (strain 323), Penicillium expansum and Purpureocillium lilacinum (73.1±2.32 mm; 68.1±2.24 mm; 62.2±3.11 mm, respectively).
When the Trichoderma is sown to the test object on the second day, the radius of T. longibrachiatum colonies is significantly larger than the radius of the colonies of test objects, reaching the highest values in the variant with Penicillium expansum, Fusarium sp. (strain 323) and Fusarium sp. (strain 376j) (70.7±2.49 mm; 69.4±1.74 mm; 69.4±1.82 mm, respectively). When the Trichoderma is introduced to the test object on the fourth day, the radius of T. longibrachiatum colonies overlaps the radius of the test objects only if Penicillium expansum and Fusarium sp. (strain 323) are used as test objects.
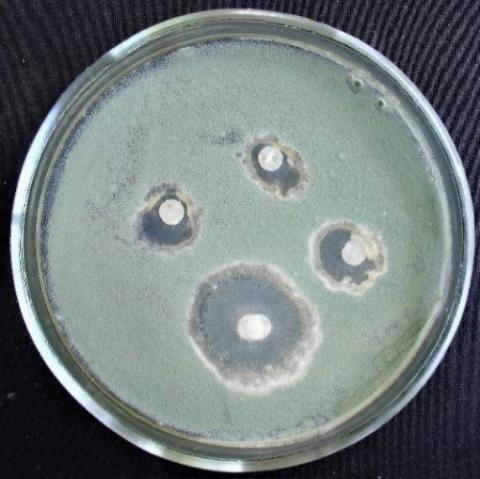
The zone of growth suppression of phytopathogenic fungi by extracts of representatives of the genus Trichoderma by the method of paper disks on day 5 ranged from 2.5±0.14 to 17.7±0.23 mm (Table 6, Figures 18-25).
The most significant growth suppression zones of test objects were noted in the variant with T. koningiopsis and T. viridescens (Figures 18-25). Notably, in some fungi, with such cultivation, sporulation occurs much later (Figure 19).
When studying the effect of Trichoderma strains on the growth of pea roots and seedlings, it was found that in the variety Nikitka the length of the aboveground part on the eighth day in all options for the experiment was less than in the control; while in the variety Zhigalova 112 the length of the aboveground part on the eighth day in almost all variants of the experiment is slightly greater than in the control (Table 7). The exception is the variant with T. viridescens.
As for the underground part of the Nikitka and Zhigalova 112 peas, its length in almost all options for the experiment exceeds that in the control. The exception again is the variant with T. viridescens on Zhigalova 112. Apparently, the strains of T. harzianum, T. koningiopsis, and T. longibrachiatum increase the growth rate of the pea root system, which is also characteristic of some strains of T. viride [11, 38]
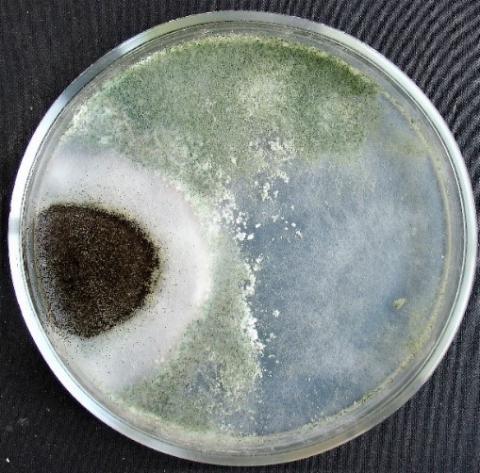
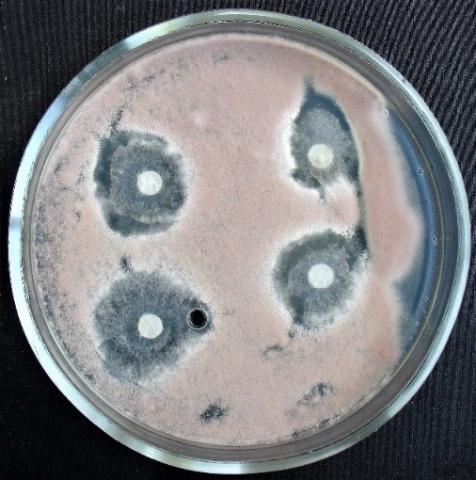
Figure 18. Alternaria sp. growth suppression zone by an extract from T. viridescens mycellium
Figure 19. Aspergillus niger growth suppression zone by an extract from T. viridescens mycelium
Figure 20. Fusarium sp. (strain 323) growth suppression zone by an extract from T. viridescens mycelium
Figure 21. Fusarium sp. (strain 323) growth suppression zone by an extract from T. viridescens mycelium
Figure 22. Penicillium expansum growth suppression zone by an extract from T. viridescens mycelium
Figure 23. Purpureocillium lilacinum growth suppression zone by an extract from T. viridescens mycelliu
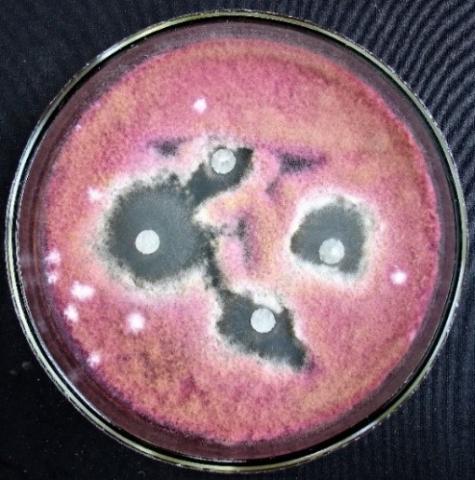
Figure 24. Fusarium sp. (strain 323) growth suppression zone by an extract from T. koningiopsis mycelium
Figure 25. Penicillium expansum growth suppression zone by an extract from T. koningiopsis mycelium
Table 7. Effect of strains of the genus Trichoderma on the growth of pea roots and seedlings, on day 8, cm
|
Test Object |
Processing Method (Experiment Variant) |
Length of the Ground Part of the Test Object |
Length of the Underground Part of the Test Object |
|
Nikita peas |
Control (water) |
6.9±0.36 |
48.8±6.07 |
|
Trichoderma viridescens |
6.0±0.42 |
67.6±6.04 |
|
|
Trichoderma koningiopsis |
6.8±0.53 |
58.0 ±5.06 |
|
|
Trichoderma harzianum |
6.8±0.48 |
58.6±2.81 |
|
|
Trichoderma longibrachiatum |
6.6±0.36 |
84.2±3.23 |
|
|
Zhigalova 112 peas |
Control (water) |
6.5±0.71 |
47.6±4.31 |
|
Trichoderma viridescens |
6.2±0.77 |
46.3±2.20 |
|
|
Trichoderma koningiopsis |
7.7±0.14 |
66.9±6.13 |
|
|
Trichoderma harzianum |
6.8±0.58 |
51.5±5.10 |
|
|
Trichoderma longibrachiatum |
6.9±0.71 |
72.2±7.62 |
The objective of the research was to extract and assess the counteractive properties of various Trichoderma strains found in the soil of Southeast Kazakhstan against widespread phytopathogenic fungi. Additionally, the study was designed to scrutinize the influence of these particular strains of Trichoderma on the development of peas, a staple crop in the region. Analysing the data obtained, it can be concluded that with simultaneous inoculation of Trichoderma and test objects on Petri dishes to determine the inhibitory activity of various species of the genus Trichoderma by the method of double cultures, the radius of T. longibrachiatum colonies significantly overlaps the radius of all test objects. Colonies of T. harzianum and T. viridescens are characterised by a smaller radius than colonies of T. longibrachiatum, but they are also larger in size than colonies of all test objects.
When analysing the influence of Trichoderma strains on the growth of pea roots and seedlings, it was found that in the variety Nikitka, the length of the aboveground part on the eighth day was less than that of the control; whereas in the variety Zhigalova 112, the length of the aboveground part on the eighth day in almost all the experimental variants was somewhat greater than that of the control; as for the underground part of the peas of Nikitka and Zhigalova 112, its length in almost all the experimental variants exceeded that of the control. The exception in both cases was the variant with T. viridescens on Zhigalova 112. All the strains obtained have a sufficiently high inhibitory activity against phytopathogenic fungi and increase the intensity of growth of the pea root system.
The findings from this study suggest a significant potential for the application of the isolated Trichoderma strains in agricultural practices. Given their demonstrated inhibitory activity against several phytopathogenic fungi, these strains could potentially be utilized as a biocontrol agent, providing a more sustainable and environmentally friendly alternative to traditional chemical fungicides. The observed enhancement of the pea root system growth also hints at the possibility of using these Trichoderma strains as bio-stimulants to promote plant development, which could potentially lead to improved crop yields. These promising results, however, are only the initial step. Understanding the precise mechanisms by which these Trichoderma strains interact with both phytopathogenic fungi and plants is crucial for their effective utilization in agriculture. Future research should, therefore, focus on elucidating these interactions at a molecular and biochemical level. Moreover, while the current study was conducted under controlled laboratory conditions, the effectiveness and impact of these strains need to be evaluated under real field conditions, considering various environmental factors and their interaction with other soil microorganisms. Thus, field trials should be included in future investigations to validate these preliminary findings.
[1] Harman, G.E., Howell, S.R., Viterbo, A., Chet, I., Lorito, M. (2004). Trichoderma species are opportunistic, avirulent plant symbionts. Nature Reviews Microbiology, 2: 43-56. https://doi.org/10.1038/nrmicro797
[2] Samuels, J.J. (2006). Trichoderma: Systematics, the sexual state, and ecology. Phytopathology, 96(2): 195-206. https://doi.org/10.1094/PHYTO-96-0195
[3] Alimova, F.K. (2005). Modern trichoderma / hypocrea system. Natural Sciences, 147: 28-55. https://cyberleninka.ru/article/n/sovremennaya-sistema-trichoderma-hypocrea.
[4] Alimova, F.K. (2006). Industrial application of fungi of the genus Trichoderma. Kazan: Kazan State University.
[5] Yakimenko, E.E. (1992). Micromycetes of forest nursery soils. Mycology and Phytopathology, 26: 480-485.
[6] Bailey, B.A., Bae, H., Strem, M.D., Roberts, D.P., Thomas, S.E., Crozier, J., Samuels, G.J., Choi, I.-Y., Holmes, K.A. (2006). Fungal and plant gene expression during the colonization of cacao seedlings by endophytic isolates of four Trichoderma species. Planta, 224: 1449-1464. https://doi.org/10.1007/s00425-006-0314-0
[7] Yu, J., Zhang, J., Wu, M., Yang, L., Chen, W., Lee, G. (2016). Screening of Trichoderma isolates based on multiple criteria for biological control of Botrytis cinerea in tomatoes. Biological Control, 101: 31-38. https://doi.org/10.1016/j.biocontrol.2016.06.006
[8] Taylor, J., Jacobson, D., Crocken, S., Kasuga, T., Geyser, D., Hibbett, D., Fisher, M. (2000). Phylogenetic recognition of species and species concepts of fungi. Fungal Genetics and Biology, 31(1): 21-32. https://doi.org/10.1006/fgbi.2000.1228
[9] Sempere, F., Santamarina, M. (2007). In vitro biocontrol analysis Alternaria alternata (Fr.) Keissler under various environmental conditions. Mycopathology, 163(3): 183-190. https://doi.org/10.1007/s11046-007-0101-x
[10] Samuels, G.J., Ismail, A. (2009). Trichoderma evansii and T. lieckfeldtiae: Two new T. hamatum-like species. Mycology, 101(1): 142-156. https://doi.org/10.3852/08-161
[11] Gneusheva, I.A., Pavlovskaya, N.E., Yakovleva, I.E. (2010). Biological activity of fungi of the genus Trichoderma and their industrial application. Bulletin of OrelGAU, 3(24): 36-39.
[12] Guigon, L.S., Guerrero, P.V., Vargas, A.F., Carvajal, M.E., Avila Quesada, G.D., Bravo, L., Ruokko, M., Lanzuis, S., Wu, S., Lorito, M. (2010). Identification Molecular de Cepas Nativas de Trichoderma ssp. Su tasa de crecimiento in vitro and Antagonismo contra Hongos. Fitopatogenos. Revista Mexicana de Fitopatología, 28(2): 87-96.
[13] Shcherbakova, T., Popushoi, I., Odobescu, V. (2010). Antagonistic activity of the fungus Trichoderma virens against the pathogens Sclerotinia sclerotiorum and Fusarium SP. Buletinul Academiei de Ştiinţe a Moldovei. Ştiinţele Vieţii, 1(310): 106-114.
[14] Alvarado-Marchena, L., Rivera-Mendes, V. (2016). Molecular identification of Trichoderma spp. in garlic and onion fields and in in vitro antagonism tests on Sclerotium cepivorum. Revista Brasileira de Ciência do Solo, 40: e0150454. https://doi.org/10.1590/18069657rbcs20150454
[15] Alexandrova, A.A., Velikanov, L.L., Sidorov, I.I. (2004). Historical overview and modern system of the genus Trichoderma. Mycology and Phytopathology, 38: 3-20.
[16] Alexandrova, A.V., Velikanov, L.L. (1999). First find of Trichoderma satumisporum in Russia. Mycology and Phytopathology, 33: 304-306.
[17] Maurer, V., Pinchuk, A. (2022). Optimisation of substrate composition and level of mineral nutrition as the basis of improving the production of decorative plants in container culture. Ukrainian Journal of Forest and Wood Science, 13(4): 64-73. https://doi.org/10.31548/forest.13(4).2022.64-73
[18] Lyubchyk, S., Shapovalova, O., Lygina, O., Oliveira, M.C., Appazov, N., Lyubchyk, A., Charmier, A.J., Lyubchik, S., Pombeiro, A.J.L. (2019). Integrated green chemical approach to the medicinal plant carpobrotus edulis processing. Scientific Reports, 9(1): 18171. https://doi.org/10.1038/s41598-019-53817-8
[19] Bazaliy, V., Domaratskiy, Ye., Pichura, V., Kozlova, O., Jarosiewicz, A. (2022). Realization of the adaptive yield potential of the assortment of winter wheat in the Steppe zone under different growing conditions. Ukrainian Black Sea Region Agrarian Science, 26(4): 30-39. https://doi.org/10.56407/2313-092X/2022-26(4)-3
[20] Biyashev, K.B., Kirkimbaeva, Z.S., Biyashev, B.K., Makbuz, A.Z., Bulegenova, M.D. (2019). Determination of the level of resistance of probiotic strain escherichia coli 64g to hydrochloric acid, bile and antimicrobial agents. Ecology, Environment and Conservation, 25(4): 1930-1933.
[21] Sadykova, V.S., Gromovykh, T.I., Kurakov, A.V., Likhachev, A.N. (2006). Prospects for the creation of new forms of biological products based on Siberian biocontrol strains of fungi of the genus Trichoderma. Bulletin of Biotechnology and Physicochemical Biology, 2: 20-21.
[22] Sadykova, V.S., Gromovykh, T.I., Likhachev, A.N., Kurakov, A.V., Ushanova, V.M. (2007). Biological activity of Siberian strains of Trichoderma as a selection criterion for the creation of new generation plant protection biological products. Biotechnology, 43: 12-17.
[23] Brazhnikova, Y.V., Shaposhnikov, A.I., Sazanova, A.L., Belimov, A.A., Mukasheva, T.D., Ignatova, L.V. (2022). Phosphate mobilization by culturable fungi and their capacity to increase soil P availability and promote barley growth. Current Microbiology, 79(8): 240. https://doi.org/10.1007/s00284-022-02926-1
[24] Rakhimova, E.V., Asylbek, A.M., Kyzmetova, L.A., Ermekova, B.D., Dzhetigenova, U.K. (2018). Species of the genus Trichoderma in the rhizosphere of the main forest-forming species of the Zailiyskiy Alatau. Higher School of Kazakhstan, 4: 110-114. https://doi.org/10.30546/2664-5297.2021.4.2.5
[25] Asylbek, A.M., Rakhimova, E.V., Sypabekkyzy, G., Urmanov, G.A., Aytymbet, Zh., Dzhunuskanova, B.E. (2020). Representatives of the genus Trichoderma in the rhizosphere of plants in Kungey Alatau (Kazakhstan). Problems of Modern Science and Education, 10(155): 22-26.
[26] Watanabe, T. (2002). Pictorial Atlas of Soil and Seed Fungi: Morphology of Cultivated Fungi and Definition. Boca Raton, CRC Press.
[27] Hassan, M.M., Farid, M.A., Gaber A. (2019). Rapid identification of Trichoderma koningiopsis and Trichoderma longibrachiatum using amplified region markers characterized by the sequence. Egyptian Journal of Biological Pest Control, 29: 13. https://doi.org/10.1186/s41938-019-0113-0
[28] Khomenko, T., Tonkha, O., Pikovska, O. (2023). Humus and nitrogen content of sod-podzolic soil under the influence of biopreparations for potato cultivation. Plant and Soil Science, 14(1): 82-95. https://doi.org/10.31548/plant1.2023.82
[29] Zakharchuk, O., Vyshnevetska, O., Ionitsoi, Ye. (2022). Seed production of winter cereals – the basis of national selection and the task of Ukrainian breeding science in 2022. Ekonomika APK, 29(2): 10-19. https://doi.org/10.32317/2221-1055.202202010
[30] Yaklich, V.M. (2009). European species of Hypocrea Part I. Green spore species. Studies in Mycology, 63: 1-91. https://doi.org/10.3114/sim.2009.63.01
[31] Yaklich, V.M. (2011). European species Hypocrea Part II. Species with green spores. Variety of Mushrooms, 48: 1-250. https://doi.org/10.1007/s13225-011-0088-y
[32] Dutbayev, Y., Kuldybayev, N., Daugaliyeva, S., Ismailova, E., Sultanova, N., Özer, G., Yessimbekova, M. (2022). Occurrence of spot blotch in spring barley caused by bipolaris sorokiniana shoem. in south-eastern kazakhstan. Scientific World Journal, 2022 https://doi.org/10.1155/2022/3602996
[33] Li, Z., Liu, Y., Zhang, Y., Shao, X., Luo, Q., Guo, X., Chong, Y. (2017). Naturally occurring resistance-associated variants to hepatitis C virus direct-acting antiviral agents in treatment-naive HCV genotype 6a-infected patients. BioMed Research International, 2017 https://doi.org/10.1155/2017/9849823
[34] Mugizi, D.R., Muradrasoli, S., Boqvist, S., Erume, J., Nasinyama, G.W., Waiswa, C., Magnusson, U. (2015). Isolation and molecular characterization of brucella isolates in cattle milk in Uganda. BioMed Research International. https://doi.org/10.1155/2015/720413
[35] GeneBank database. (2021). http://www.ncbi.nih.gov, accessed on Dec. 5, 2021.
[36] Ignatova, L., Kistaubayeva, A., Brazhnikova, Y., Omirbekova, A., Mukasheva, T., Savitskaya, I., Karpenyuk, T., Goncharova, A., Egamberdieva, D., Sokolov, A. (2021). Characterization of cadmium-tolerant endophytic fungi isolated from soybean (Glycine max) and barley (Hordeum vulgare). Heliyon, 7(11): e08240. https://doi.org/10.1016/j.heliyon.2021.e08240
[37] Mero, G., Skenderasi, B., Shahini, E., Shahini, S., Shahini, E. (2023). Main directions of plants integrated protection in the conditions of organic agriculture. Scientific Horizons, 26(3): 101-111. https://doi.org/10.48077/SCIHOR3.2023.101
[38] Turbekova, A., Oshergina, I., Kurmangozhinov, A., Ten, E., Amantayev, B., Kipshakbaeva, G. (2022). Evaluation of the genetic material of the grass pea (Lathyrus Sativus L.) seed collection in Northern Kazakhstan. OnLine Journal of Biological Sciences, 22(2): 191-200. https://doi.org/10.3844/ojbsci.2022.191.20