Xiurong Si* | Ding Ding | Jianhua Zhou | Zhangwei Cao
© 2020 IIETA. This article is published by IIETA and is licensed under the CC BY 4.0 license (http://creativecommons.org/licenses/by/4.0/).
OPEN ACCESS
This paper mainly investigates the inhibitory effect and mechanism of vanillin on mixed-culture biofilm formation, and discusses the feasibility of controlling membrane fouling with vanillin. The biofilms were cultured under static conditions and a microfluidic chamber, respectively. The results show that vanillin can effectively inhibit the microbial adhesion and biofilm formation; the inhibition rate increases with the vanillin concentration. After 12h treatment with 300mg/L vanillin, the inhibition rate of biofilm formation reached 85.34%. Besides, vanillin can mitigate bacterial adhesion by reducing the microbial secretion of exoprotein and exopolysaccharide, exhibit an inhibitory effect on the expression of N-acyl-l-homoserine lactones (AHLs), but had no effect on environmental deoxyribonucleic acid (eDNA). After being treated with 300mg/L vanillin, exoprotein, exopolysaccharide and AHLs decreased by 28.48%, 17.23% and 46.64%, respectively. Furthermore, there is a positive correlation between the AHLs and the content of extracellular polymeric substances (EPSs); the AHLs-mediated quorum sensing (QS) might be related to microbial adhesion through the regulation of EPS. Finally, vanillin has a great potential in membrane fouling control. The research findings provide a good reference for biofilm control in water purification and wastewater treatment systems.
vanillin, biofilms, multi-species, inhibition rate
Biofilm is a highly structured microbial community formed by microorganisms attached to the surface of inert or active solid materials, as well as the extracellular polymeric substances (EPSs) secreted by the microorganisms themselves [1]. The presence of biofilm could induce a series of environmental problems, namely, membrane fouling, pipe blockage, metal corrosion, and reduced disinfection efficiency [2, 3]. Therefore, many bactericides or antibacterial agents have been used to control biofilms [4, 5]. However, the release of these substances brings new environmental risks, and increases the resistance of bacteria [6]. What is worse, the bactericidal effect of these substances is suppressed by the three-dimensional (3D) structure formed by the high density of bacteria and the huge number of EPSs [7]. It has been reported that microorganisms in the aggregated state are 1,000 times more resistant than those in the suspended state to traditional bactericides or antibacterial agents [8]. Therefore, there is an urgent need for a non-microbicidal method to control biofilms.
Many researchers have attempted to regulate the biofilm development with small modules that inhibit microbial activity. The microbial regulation of biofilm formation is achieved through quorum sensing (QS) [9]. There are three types of QS systems: the QS mediated by N-acyl-l-homoserine lactones (AHLs) as signal molecules in Gram-negative bacteria; the QS mediated by modified autoinducing oligopeptides (AIPs) as signal molecules in Gram-positive bacteria; and the non-specific languages for interspecific communication mediated by AI-2. To further inhibit biofilm formation, QS inhibitors (QSIs) could be used to interfere with the QS, and cut off the communication of signal molecules, reducing the risk of bacterial resistance.
In wastewater treatment, the interference and blocking of QS are mainly concentrated in the pollution control of membrane modules in membrane bioreactor (MBR) and reverse osmosis (RO) systems [10, 11]. Vanillin (4-hydroxy-3-methoxybenzaldehyde) is a chemical substance proven as a promising QSI by interference with AHL receptors. There has been reports about the inhibition of vanillin on pure bacterial biofilms [12]. Nevertheless, few scholars have explored the inhibition of vanillin on mixed-culture biofilms. In the real-world, most biofilms are composed of multi-species culture. Therefore, it is necessary to study the inhibitory effect of vanillin on the biofilms of multi-species culture.
Through the above analysis, this paper investigates the inhibitory effect and mechanism of vanillin on the biofilm formation of mixed culture, and discusses the feasibility of controlling membrane fouling with vanillin. The research results shed important new light on biofilm formation and biofouling control under non-microbicidal mechanisms.
2.1 Chemical reagents and microorganisms
Vanillin was purchased from Sangon Biotech (Shanghai, China). The mixed culture was obtained from the activated sludge of a municipal wastewater treatment plant (Beijing, China), and analyzed by a MiSeq system of Majorbio (Shanghai, China).
The activated sludge was washed three times with phosphate buffer solution (PBS) to remove EPSs and organic matters in the sludge solution, and resuspended in synthetic wastewater to prepare a mixed bacterial suspension (100 mg VSS/L) (approximately 108 CFU/mL). The synthetic wastewater contains (mg/L): chemical oxygen demand (COD, calculated as glucose) 825; NH4Cl 192; KH2PO4 35.1; NaCl 100; MgSO4 100; CaCl2 10; pH = 6.5-7.5.
2.2 Effect of vanillin on biofilm formation of the mixed culture
The bacterial suspensions containing one of the five concentrations of vanillin (0, 50, 100, 150, and 300 mg/L) were added to 12-well plates of polystyrene with 4mL per well. After that, the 12-well plates were incubated for 12h and 24h, respectively, at a constant temperature of 30°C. The amount of biofilm was quantified through crystal violet staining [13].
To mimic the biofilm formation in actual wastewater treatment, a microfluidic chamber was taken as the biofilm formation device, and a glass slide was selected as a biofilm adhesion carrier, aiming to reveal the effect of vanillin on biofilm formation. The bacterial suspensions were prepared as above. Two of the channels were fed with 300mg/L vanillin, and the other two served as controls at a flow rate of 2mL/min. After 1 day and 7 days of operation, the slides were taken out from the microfluidic chamber, and the amount of attached biofilm was measured through crystal violet staining. Finally, the stained slides were placed under a microscope to observe the adhesion of microorganisms at different positions.
2.3 The effect of vanillin on the adhesion of mixed culture on different substrate surfaces
The inhibitory effect on the adhesion features of microorganisms depends heavily on the surface features of the attached substrate. Therefore, this paper further explores the effect of vanillin on the adhesion of microorganisms on the surface of three substrates with different hydrophilic features: polystyrene (6-well plate), glass sheet, and polymethyl methacrylate. The contact angles of the three substrates were set to 93.4°, 22.0°, and 81.9°, respectively. The sludge suspension, substrate concentration, and operating conditions were kept the same as those of the biofilm formation experiments above. After incubation for 24h, the substrate surfaces were dried, stained, and imaged with an Olympus camera.
2.4 The mitigation effect of vanillin on membrane fouling
The mitigation effect of vanillin on membrane fouling was investigated through the following operations: polyamide flat membranes with a diameter of 3.5cm were placed in 6-well culture plates. Then, 10mL mixed culture solution and a particular amount of vanillin were added to each plate to attain desired concentrations (50-300mg/L). Next, these plates were statically incubated at 30 ℃ for 12h to form biofilms. After incubation, each membrane was placed in 15mL PBS, and sonicated for 15min to remove the attached microorganisms. After 15s mixing, the absorbance of suspension was determined at 600nm. The bacterial optical density was measured directly, due to the combination of crystal violet dye and the membrane material.
2.5 The effect of vanillin on the distribution of microbial EPSs
The effect of vanillin on the distribution of microbial EPSs was examined to further showcase its inhibition of biofilm formation. First, a certain concentration of sludge suspension and substrate described as above were put into a 50mL centrifuge tube, and vanillin was added to make the final concentration 0 and 300mg/L, respectively. The mixture was incubated at 30 ℃ for 12h to determine the change in EPS content. The EPSs were extracted and measured by the method of Wang et al. [14].
2.6 The effect of vanillin on the distribution of microbial AHLs
The method of Lv et al. [15] was slightly modified to detect the AHLs. First, the sludge was centrifuged at 8,000g for 5min, resuspended in 5mL sterile water, and then sonicated at 20kHz for 30min. Then, the suspension was filtered through a 0.22μm membrane to obtain the test sample. The reported strains NTL4 were inoculated into minimal medium (MM), supplemented with 50μg/mL gentamicin, and then cultured at 28 ℃. The bacterial culture was diluted to a OD600 value of 0.1 with an antibiotic-free MM, and was prepared in 12-well plates. These plates were incubated for 16-18h at 28℃,180rpm. Then, 200μL 20mg/ml X-gal was added, and shaken for 120min. The absorbance of the suspension was determined at 635nm. The ratio of sample to blank sample OD635 was defined as the relative content of AHLs.
3.1 The effect of vanillin on biofilm formation of mixed culture
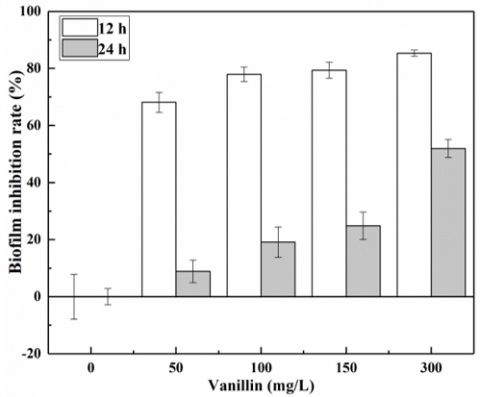
Figure 1 shows the effect of vanillin on biofilm formation of mixed culture. After the microorganisms were treated with 50~300mg/L vanillin for 12h, the adhesion decreased by 68.13%~85.34%; after 24h, the adhesion was reduced by 8.89%~51.96%. It can be seen that the inhibition rate increased with vanillin concentration, but decreased with the elapse of time.
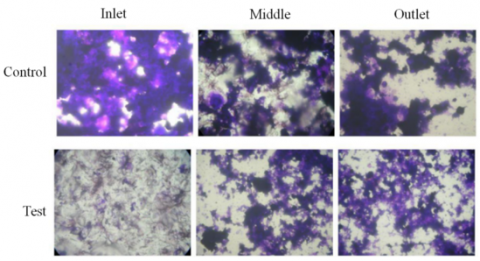
The effect of vanillin on biofilm formation was also investigated at different flow conditions. After the microfluidic chamber had been operating for 24h, the inhibition rate of 300mg/L vanillin on microbial adhesion was 57.76%. Three points were selected for observation at the inlet, middle, and outlet of the microfluidic chamber, respectively. The observation results in Figure 2 show that color of crystal violet in the control group was much darker than that in the test group, indicating that more microorganisms were attached to the slide surface of the control group, which was obtained at the three observation points.
Next, the microfluidic chamber operated for 7 more days to better reveal the effect of vanillin on microbial adhesion under long-term water conservancy conditions. The inhibition rate of vanillin on microbial adhesion dropped to 39.51%. This means vanillin has a high inhibition rate on biofilm formation, whether it is cultured under static conditions or in a microfluidic chamber; however, the inhibition rate will decline with the elapse of time. A possible reason lies in the changing composition of the extracellular matrix in the later phase of biofilm formation [16]. Rice et al. [17] also found that the deoxyribonuclease has relatively strong inhibition of S. aureus biofilm formation in the early phase than in the later phase. Lv et al. [15] studied the effects of vanillin and proteinase K on the adhesion features of aerobic granular sludge, and discovered that inhibitory effect of these two substances on the particle adhesion ability is stronger after 12h treatment than after 24h treatment.
Figure 1. Inhibitory effect of vanillin on biofilm formation at different exposure time
Figure 2. Effect of vanillin on biofilm formation at different flow conditions (microscopic images)
3.2 The effect of vanillin on the adhesion of mixed culture on different substrate surfaces
As the first step in biofilm formation, the microbial adhesion to the surface of the carrier could occur through specific or non-specific bacterial-surface interactions [18]. Some surface features can affect the adhesion of microorganisms, as well as the effect of some inhibitors. Therefore, this paper probes into the effect of vanillin on microbial adhesion on different substrate surfaces. In the presence of vanillin, the biomass attached to the substrate surfaces was recorded through crystal violet staining
As shown in Figure 3, whether it is on the surface of hydrophobic polystyrene or hydrophilic polymethyl methacrylate and glass, vanillin always exhibited a good inhibitory effect on microbial adhesion. In other words, vanillin has a great prospect in biofilm control on the surface of different materials.
Figure 3. Inhibitory effect of vanillin on biofilm formation on different substrate surfaces
3.3 The mitigation effect of vanillin on membrane fouling
The microbial adhesion on the membrane surface was further studied to explore the possibility of vanillin on biofouling control on membrane surface. Figure 4 shows the amounts of microorganisms attached to the polyamide membrane surface at different concentrations of vanillin. It can be seen that vanillin treatment greatly weakened the microbial adhesion on the membrane surface. For instance, the amount of microorganisms attached to the membrane surface decreased by 86.12% after being treated with 300mg/L vanillin.
Figure 4. Effect of vanillin on microbial adhesion of mixed culture on the surface of polyamide membrane
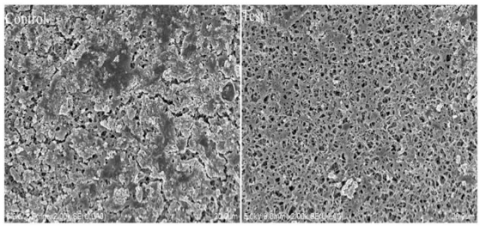
This finding was confirmed by the scanning electron microscope (SEM) images (Figure 5). In the control group, the microbial community formed on the membrane surface was relatively dense, and the biofilms were connected into pieces. By contrast, in the test group, the microorganisms attached to the membrane surface were quite sparse, leaving lots of blank areas; the amount of microorganisms was much fewer than that in the control group. Hence, vanillin has the potential to be applied in biofouling control on membrane surface.
3.4 The effect of vanillin on the secretion of microbial EPSs and AHLs
The microbial EPSs and AHLs play important roles in the microbial adhesion in the early phase and the biofilm growth in the later phase. Therefore, the microbial EPSs (e.g. exoproteins, exopolysaccharides, and environmental deoxyribonucleic acid (eDNA)) and AHLs were measured to further explore the inhibitory mechanism of vanillin on biofilm formation.
Figure 5. Effect of vanillin on microbial adhesion of mixed culture on the surface of polyamide membrane (SEM images)
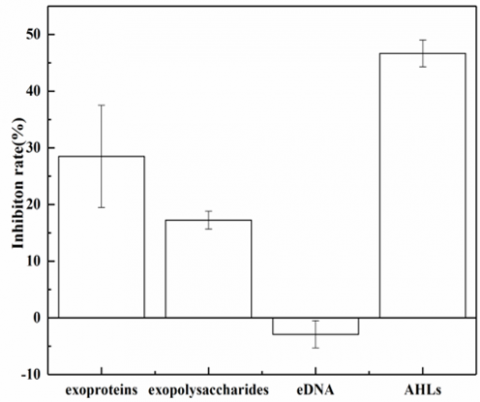
Figure 6 presents the effect of vanillin on the production of EPSs and AHLs in suspended microorganisms. It can be seen that the exoproteins, exopolysaccharides, and AHLs in the test group decreased by 28.48%, 17.23% and 46.64%, respectively, from the levels of the control group. However, the eDNA content of the test group was basically the same as that in the control group. These results indicate that vanillin has a certain inhibitory effect on the secretion of exoproteins, exopolysaccharides, and AHLs in the microorganisms, but has no effect on the production of eDNA.
Figure 6. The effect of vanillin on the production of EPSs and AHLs
Biofilm formation and QS are often used to describe the social behaviors of microorganisms. Studies have shown a close relationship between QS and biofilms. Besides participating in biofilm formation, the QS has a direct bearing on biofilm detachment [19, 20]. Katebian et al. [21] and Ham et al. [22] demonstrated that QSIs have the potential to weaken membrane biofouling.
Exoproteins and exopolysaccharides, as the main components of EPSs, provide strong support to biofilm formation. If the EPS content is insufficient, or if changes occur to the ratio between exoproteins-exopolysaccharides, the spatial architecture of EPSs will be destroyed [13], halting biofilm formation or causing biofilms to disintegrate. In addition, there are reports on the role of the QS in in regulating EPS production in pure culture systems [23-25].
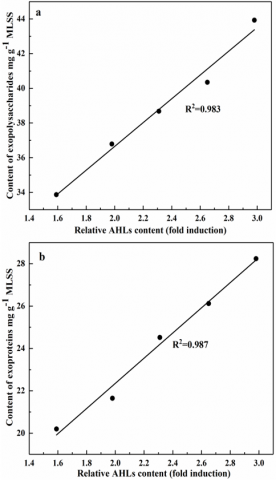
The results of our research suggest a positive correlation between the EPS content in the activated sludge of mixed culture and the QS activity mediated by the AHLs signal molecule (Figure 7). It can be speculated that AHLs-mediated QS is involved in microbial adhesion of mixed culture through the regulation of exoproteins and exopolysaccharides. This finding echoes with the conclusions of Yeon et al. [10] and Jiang et al. [11]: the AHLs quenching enzyme can reduce the EPSs in the MBR reactor and sludge system.
Figure 7. The relationship between exopolysaccharides (a), exoproteins (b), and relative AHLs content
This paper mainly investigates the inhibitory effect and mechanism of vanillin on mixed-culture biofilm formation, and establishes a novel biofilm control strategy based on interfering QS systems. The main conclusions are as follows:
(1) Vanillin can effectively inhibit microbial adhesion and biofilm formation, whether it is cultured under static conditions or in a microfluidic chamber. The inhibition rate increases with vanillin concentration.
(2) Vanillin can mitigate bacterial adhesion by reducing the microbial secretion of exoprotein and exopolysaccharide, and exhibit a certain inhibitory effect on the expression of AHLs signal molecules, but has no effect on eDNA.
(3) There is a positive correlation between the EPS content and AHLs. The AHLs-mediated QS might be related to microbial adhesion through the regulation of EPS.
(4) Vanillin boasts a great potential in environmental membrane fouling control.
This research was supported by the National Natural Science Foundation of China (Grant No.: 51808008), “Three Three Three” Talents Project of Heibei Province (A202001024), the Natural Science Foundation of Hebei Province (Grant No.: E2017409004) and Hebei Collaborative Innovation Center for Aerospace Remote Sensing Information Processing and Application.
[1] Shi, Y., Huang, J., Zeng, G., Gu, Y., Chen, Y., Hu, Y., Tang, B., Zhou, J., Yang, Y., Shi, L. (2017). Exploiting extracellular polymeric substances (EPS) controlling strategies for performance enhancement of biological wastewater treatments: an overview. Chemosphere, 180: 396-411. https://doi.org/10.1016/j.chemosphere.2017.04.042
[2] Tarifa, M.C., Genovese, D., Lozano, J.E., Brugnoni, L.I. (2018). In situ microstructure and rheological behavior of yeast biofilms from the juice processing industries. Biofouling, 34(1): 74-85. https://doi.org/10.1080/08927014.2017.1407758
[3] Jeong, S., Cho, K., Jeong, D., Lee, S., Leiknes, T., Vigneswaran, S., Bae, H. (2017). Effect of engineered environment on microbial community structure in biofilter and biofilm on reverse osmosis membrane. Water Research, 124: 227-237. https://doi.org/10.1016/j.watres.2017.07.064
[4] Kang, J., Dietz, M.J., Li, B. (2019). Antimicrobial peptide LL-37 is bactericidal against Staphylococcus aureus biofilms. PloS One, 14(6): e0216676. https://doi.org/10.1371/journal.pone.0216676
[5] Rakesh K.P., Vivek H.K., Manukumar H.M., Shantharam C.S., Bukhari S.N.A., Qin H.L., Sridhara, M.B. (2018). Promising bactericidal approach of dihydrazone analogues against bio-film forming Gram-negative bacteria and molecular mechanistic studies. RSC Advances, 8(10): 5473-5483. https://doi.org/10.1039/C7RA13661G
[6] Xu, N., Cheng, H., Xu, J., Li, F., Gao, B., Li, Z., Gao, C., Huo, K., Fu, J., Xiong, W. (2017). Silver-loaded nanotubular structures enhanced bactericidal efficiency of antibiotics with synergistic effect in vitro and in vivo. International journal of Nanomedicine, 12: 731-743. https://doi.org/10.2147/IJN.S123648
[7] Sharma, G., Sharma, S., Sharma, P., Chandola, D., Dang, S., Gupta, S., Gabrani, R. (2016). Escherichia coli biofilm: development and therapeutic strategies. J Appl Microbiol. 121(2): 309-319. https://doi.org/10.1111/jam.13078
[8] Rasmussen T.B., Givskov M. (2006). Quorum-sensing inhibitors as anti-pathogenic drugs. International Journal of Medical Microbiology, 296(2-3): 149-161. https://doi.org/10.1016/j.ijmm.2006.02.005
[9] Feng, L., Wu, Z., Yu, X. (2013). Quorum sensing in water and wastewater treatment biofilms. Journal of Environmental Biology, 34(2S): 437-444.
[10] Yeon, K.M., Cheong, W.S., Oh, H.S., Lee, W.N., Hwang, B.K., Lee, C.H., Beyenal, H., Lewandowski, Z. (2009). Quorum sensing: a new biofouling control paradigm in a membrane bioreactor for advanced wastewater treatment. Environmental science & technology, 43(2): 380-385. https://doi.org/10.1021/es8019275
[11] Jiang, W., Xia, S.Q., Liang, J., Zhang, Z.Q., Hermanowicz, S.W. (2013). Effect of quorum quenching on the reactor performance, biofouling and biomass characteristics in membrane bioreactors. Water research, 47(1): 187-196. https://doi.org/10.1016/j.watres.2012.09.050
[12] Ponnusamy, K., Paul, D., Kweon, J.H. (2009). Inhibition of quorum sensing mechanism and Aeromonas hydrophila biofilm formation by vanillin. Environmental Engineering Science, 26(8): 1359-1363. https://doi.org/10.1089/ees.2008.0415
[13] Xiong, Y., Liu, Y. (2013). Importance of extracellular proteins in maintaining structural integrity of aerobic granules. Colloids and Surfaces B: Biointerfaces, 112: 435-440. https://doi.org/10.1016/j.colsurfb.2013.07.060
[14] Wang, X., Zhang, B., Shen, Z., Qiu, Z., Chen, Z., Jin, M., Li, J., Wang, J. (2010). The EPS characteristics of sludge in an aerobic granule membrane bioreactor. Bioresource Technol. 101(21): 8046-8050. https://doi.org/10.1016/j.biortech.2010.05.074
[15] Lv, J., Wang, Y., Zhong, C., Li, Y., Hao, W., Zhu, J. (2014). The effect of quorum sensing and extracellular proteins on the microbial attachment of aerobic granular activated sludge. Bioresource Technology, 152: 53-58. https://doi.org/10.1016/j.biortech.2013.10.097
[16] Thomas, V.C., Thurlow, L.R., Boyle, D., Hancock, L.E. (2008). Regulation of autolysis-dependent extracellular DNA release by Enterococcus faecalis extracellular proteases influences biofilm development. Journal of Bacteriology, 190(16): 5690-5698. https://doi.org/10.1128/JB.00314-08
[17] Rice K.C., Mann E.E., Endres, J.L., Weiss E.C., Cassat J.E., Smeltzer M.S., Bayles, K.W. (2007). The cidA murein hydrolase regulator contributes to DNA release and biofilm development in Staphylococcus aureus. Proceedings of the National Academy of Sciences, 104(19): https://doi.org/10.1073/pnas.0610226104
[18] Habimana, O., Semião, A.J.C., Casey, E. (2014). The role of cell-surface interactions in bacterial initial adhesion and consequent biofilm formation on nanofiltration/reverse osmosis membranes. Journal of Membrane Science, 454: 82-96. https://doi.org/10.1016/j.memsci.2013.11.043
[19] Hu, H., He, J., Liu, J., Yu, H., Tang, J., Zhang, J. (2016). Role of N-acyl-homoserine lactone (AHL) based quorum sensing on biofilm formation on packing media in wastewater treatment process. Rsc Advances, 6(14): 11128-11139. https://doi.org/10.1039/C5RA23466B
[20] Kim, S.K., Lee, J.H. (2016). Biofilm dispersion in Pseudomonas aeruginosa. Journal of Microbiology, 54(2): 71-85. https://doi.org/10.1007/s12275-016-5528-7
[21] Katebian, L., Gomez, E., Skillman, L., Li, D., Ho, G., Jiang, S.C. (2016). Inhibiting quorum sensing pathways to mitigate seawater desalination RO membrane biofouling. Desalination, 393: 135-143. https://doi.org/10.1016/j.desal.2016.01.013
[22] Ham, S.Y., Kim, H.S., Jang, YS.., Sun, P.F., Park, J.H., Lee, J.S., Byun, Y., Park, H.D. (2019). Control of membrane biofouling by 6-gingerol analogs: Quorum sensing inhibition. Fuel, 250: 79-87. https://doi.org/10.1016/j.fuel.2019.03.145
[23] Qian, G., Zhou, Y., Zhao, Y., Song, Z., Wang, S., Fan, J., Hu, B., Venturi V., Liu, F. (2013). Proteomic analysis reveals novel extracellular virulence-associated proteins and functions regulated by the diffusible signal factor (DSF) in Xanthomonas oryzae pvoryzicola. Journal of Proteome Research, 12(7): 3327-3341. https://doi.org/10.1021/pr4001543
[24] Marketon, M.M., Glenn, S.A., Eberhard, A., González, J.E. (2003). Quorum sensing controls exopolysaccharide production in Sinorhizobium meliloti. Journal of Bacteriology, 185(1): 325-331. https://doi.org/10.1128/JB.185.1.325-331.2003
[25] Lee, K.J., Kim, J.A., Hwang, W., Park, S.J., Lee, K.H. (2013). Role of capsular polysaccharide (CPS) in biofilm formation and regulation of CPS production by quorum‐sensing in Vibrio vulnificus. Molecular Microbiology, 90(4): 841-857. https://doi.org/10.1111/mmi.12401