Ainash Nauanova![]() | Serik Shaikhin
| Serik Shaikhin![]() | Saule Ospanova
| Saule Ospanova![]() | Meruyert Makenova
| Meruyert Makenova![]() | Nazymgul Shumenova
| Nazymgul Shumenova![]() | Makpal Bostubayeva*
| Makpal Bostubayeva*![]()
© 2024 The authors. This article is published by IIETA and is licensed under the CC BY 4.0 license (http://creativecommons.org/licenses/by/4.0/).
OPEN ACCESS
Barley is one of the most important grain crops grown in all agricultural regions of the world. It is unique in its chemical composition and health benefits. In Kazakhstan, the largest country in Central Asia, barley is the second most important grain commodity after wheat. The main goal of the project was a comparative study of the effect of four local biofertilizers in the form of consortia of indigenous soil microorganisms with PGPR and PGPF properties on the grain yield of spring barley variety “Tselinny 2005” in the conditions of the Kazakhstan semi-arid steppe zone. Employing a systematic field trial design, each biofertilizer's impact was assessed through its application rates and methods, comparing against control plots without biofertilizer treatment. Grain yield was meticulously measured post-harvest, accounting for variations in environmental conditions, to ascertain the biofertilizers' contributions to crop productivity. Based on the results obtained, recommend the most effective biofertilizers to barley producers. Laboratory tests of germination energy, germination of spring barley seeds inoculated with biofertilizers, and post-embryonic development of roots and shoots showed the effective colonization potential of at least three tested biofertilizers. Single-factor field experiments over two years showed that the significant benefits of employing biofertilizers B1 and B4, which not only promote a 50% increase in spring barley grain yield but also present a sustainable and environmentally beneficial alternative to synthetic fertilizers, pesticides, and fungicides to increase barley grain yield in areas subject to abiotic and biotic stress. Utilizing these biofertilizers could reduce environmental impact, and lower production costs, offering a holistic approach to enhancing agricultural productivity in semi-arid regions.
biofertilizer, biotic and abiotic stress, colonization potential, grain yield, spring barley, crop yield
Barley is one of the most important grain crops grown in all agricultural regions of the world. It is gaining popularity among agricultural and culinary experts due to its unique chemical composition and health benefits. The publications highlight the health benefits of barley, such as lowering cholesterol and blood sugar, anti-cancer activity, antioxidant and detoxifying properties, and anti-inflammatory and anti-arthritic properties [1].
In terms of area planted and production, barley is mainly used for animal feed, brewing malt, and human food. In Kazakhstan, the largest country in Central Asia, barley is the second most important grain commodity after wheat [2]. Modern agriculture in Kazakhstan is carried out in an extensive manner and is accompanied by a noticeable decrease in soil fertility, biological and agrochemical properties, which is the main consequence of insufficient fertilization [3]. Another problem in an agricultural region, for example, Northern Kazakhstan, is the uneven condition of crops. Precipitation falls unevenly during the summer, with rain often falling in stripes. The greatest manifestation of drought is observed in areas located in the southern black soil zones. The specific yield of barley is at the level of 9.0-13.0 c/ha [4].
Currently, plant growth-promoting rhizobacteria (PGPR) are being used to reduce the negative effects of drought stress on plant growth. The use of these beneficial microbes as biofertilizers can serve to replace and/or minimize the use of chemical fertilizers, which often have negative impacts on the environment. In this regard, biofertilizers are coming into practice as an alternative and environmentally friendly method of sustainable agriculture.
Cultivated plants grown on farms in Northern Kazakhstan inevitably face biotic and abiotic stresses of water shortage during the growing season. In the research of this project, the size of the spring barley harvest was noticeably influenced by unstable agrometeorological conditions in 2021 and 2022.
In scientific periodicals, Kazakhstani studies on the effect of biofertilizers on the yield of spring barley are represented mainly by preparations from Russia [5].
The project's objective was to assess the impact of four local biofertilizers, each a consortium of native soil bacteria, on the yield of 'Tselinny 2005' spring barley in Kazakhstan's semi-arid steppe.
In this context, the goal was to optimize crop yields, and soil resources and minimize disease.
By achieving these goals, the research sought to provide practical recommendations for agricultural producers, aiming for optimized agricultural productivity and sustainable soil management practices in the challenging semi-arid conditions.
In our research, we conducted a comparative study utilizing four local biofertilizers. This focused approach allowed us to directly assess their effects on the yield of spring barley variety 'Tselinny 2005' under the semi-arid conditions of Northern Kazakhstan.
2.1 Origin and preparation of biofertilizers
Strains of microorganisms for the construction of consortia of biofertilizers B1-B4 (Table 1) were purchased from the museum collection of the Department of Soil Science and Agrochemistry at the Kazakh Agrotechnical University named after Saken Seifullin. Strains were stored in tubes on agar slate in a refrigerator at 8℃.
In selecting biofertilizers B1-B4 for our study, we focused on those with a proven track record of enhancing soil fertility and plant health in semi-arid conditions similar to Northern Kazakhstan. These biofertilizers were chosen based on their unique consortia of indigenous soil bacteria, known for their ability to fix nitrogen, solubilize phosphate, and promote root growth.
Table 1. Biofertilizers used in research
|
Biofertilizer |
Members of the Biofertilizer Consortium |
Consortium Member’s Activities |
Source of Isolation of Consortium Members |
|
|
Symbol |
Name |
|||
|
B1 |
Compo-MIX |
Streptomyces sindenensis PM9, Streptomyces griseus PM25, Bacillus aryabhattai PM62, Bacillus aryabhattai PM68, Bacillus aryabhattai PM69, Bacillus megaterium PM80B, Lentzea violacea PM86B |
Growth-stimulating, nitrogen-fixing, cellulose-destroying and fungicidal activity. |
Soils, animal and poultry waste in the farms of the Akmola region of Northern Kazakhstan. |
|
B2 |
Agrarka |
Streptomyces xantholiticus 7, Streptomyces microsporus 12, Streptomyces sioyaensis 41 |
Producing a complex of biologically active substances with fungicidal and growth-stimulating activity. |
Soils of North Kazakhstan. |
|
B3 |
Agro-MIX |
Bacillus spp., Saccharomyces spp., Acetobacter spp., Streptomyces spp. |
Growth-stimulating, nitrogen-fixing activity, and antagonism against putrefactive pathogens. |
Bran and seed surface in grain farms of Northern Kazakhstan. |
|
B4 |
Trichodermin-KZ |
The most promising fungi strains of the genus Trichoderma T134, T115, T200, identified as Tr. lignorum and Tr. album |
High antagonistic and hyperparasitic properties. |
Soils of North Kazakhstan. |
To restore the viability and metabolic activity of the target microorganisms, isolates were passaged onto prepared agar slants with a suitable medium for the microorganisms. Liquid inoculum cultures were seeded into 100 mL flasks. The resulting liquid cultures were incubated at 28℃ on a shaker at 70 rpm for 72-120 hours, depending on the biological characteristics of the microorganisms, and the titer was set at 109 CFU/ml. The resulting liquid cultures were incubated at 28℃ on a shaker at 70 rpm for 72-120 hours, based on the microorganisms' biological characteristics, setting the titer at 109 CFU/ml, where 'titer' denotes the solution's concentration, and 'CFU/ml' measures the viable microbial cells per milliliter capable of forming colonies. Suspensions of strains were taken in equal proportions and thoroughly mixed on a shaker.
Strains of Trichoderma spp. from the collection were grown in Petri dishes on a Czapek-Dox medium (pH 6.0-6.5). Next, the 7-day culture was seeded into 250 ml flasks with liquid Czapek-Dox medium and incubated at 25℃ on a shaker at 70 rpm for 7-10 days until the titer reached 109 CFU/ml. Suspensions of strains were taken in equal proportions and thoroughly mixed on a shaker.
Working solutions of biofertilizers were prepared, and to treat 5 kg of barley seeds, it was enough to prepare 1 liter of working solution of biofertilizer at the rate of 100 ml of a suspension of a consortium of microorganisms per 900 ml of sterile water.
2.2 Laboratory experiments
The study was carried out according to the standard method [6] with minor modifications. The surface of barley seeds was disinfected by immersing once for 3 minutes in a 70% ethanol solution, then for 3 minutes in a 5% sodium hypochlorite solution and washed five times by immersing in renewed sterilized distilled water, followed by a 2-hour soak in sterilized distilled water.
Concentrations of 70% ethanol and 5% sodium hypochlorite were selected based on their effectiveness in sterilization and disinfection processes in accordance with standard laboratory practice. These concentrations are effective in eliminating surface contaminants and pathogens on seeds without harming seed viability, ensuring the integrity of our experiments. This approach is widely used in agronomic research on seed treatment prior to germination testing.
After that the seeds were soaked for 2 hours in a working biofertilizer solution containing 108 cells of each bacterial strain and 108 spores of each Trichoderma strain in one ml of solution. The bottom of the Petri dishes (9 cm in diameter) was covered with sterile filter paper and irrigated with 5 ml of sterilized water. Then ten barley seeds were placed on filter paper in each Petri dish. For the control and variants of samples with biofertilizers, five Petri dishes were used for each sample variant. All plates were incubated at 24℃ for 7 days in the dark. In the control experiment, the seeds were soaked for 2 hours in sterilized distilled water and incubated in the same way as in the sample variants.
On the second day, germination energy was assessed by observing and counting the number of typical seedlings under controlled conditions of constant temperature (24℃) and high humidity. For accurate assessment of germination energy, a typical seedling was defined based on vigor, uniformity, and the absence of disease or deformity by the according to ISTA (International Seed Testing Association) 2004 recommendations [7]. The relative germination of seeds was determined by incubation in Petri dishes for five days at a constant temperature (24℃) and high humidity.
Biometric parameters of seedlings were measured on the 7th day after the start of bioinoculation. To measure the length of shoots, 20 seedlings were selected from each experimental sample with biofertilizer and the control variant.
The length of the shoot was measured with a caliper from the beginning of the curved arc of the hypocotyl knee to the cotyledons of the leaves. The term of hypocotyl knee is used to describe the curved part of a seedling's stem just above the root. Then the average shoot length was calculated for each experimental variant and the control group. The root length from the root tip to the bend of the hypocotyl curve was measured in a similar manner.
2.3 Field experiments
Field experiments were carried out in the village of Nauchny, Akmola region, located approximately at 51°37'26.9 north latitude and 71°00'57.1 east longitude. The steppe zone where the village is located is characterized by a sharply continental climate, with cold winters and dry, hot summers. The average annual temperature in the area is 1.7℃ and the average annual rainfall is recorded at 325.6 mm.
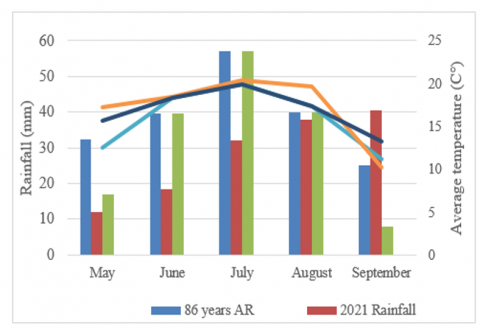
Figure 1. Meteorological conditions of the research period (Data from the Shortandy 1 meteorological station, 2021-2022)
During the barley growing season (May - September) for 2021, the sum of active temperatures was 3934℃, for 2022 3841℃, in 2021 the amount of precipitation was 132 mm, and in 2022 - 125 mm, the hydrothermal coefficient was at the level of 0.32 -0.34, which characterizes conditions in the form of a dry zone for crop growth and development (Figure 1).
Analyzing the moisture supply of the crop from June to August, one can note a lack of precipitation, especially in June-July 2021 and in May-June 2022, which is on average two times lower than the norm for long-term data. On July 29-30, 2022, heavy precipitation in the form of heavy rain was observed.
2.4 Field experiments design
Seeds of spring barley variety “Tselinny 2005” were purchased at the Research and Production Center for Grain Farming named after A.I. Baraev (NPCZH named after A.I. Baraev). This variety is certified and suitable for sowing in the Northern regions of Kazakhstan. The variety is characterized as mid-season and moderately resistant to lodging. The experimental plot was organized as a divided plot with randomized placement of options; it consisted of 5 options with 5 replications.
Randomization was achieved through a randomized complete block design (RCBD), ensuring that each biofertilizer treatment was equally represented across different plots to minimize bias and environmental variability. The plot sizes were determined based on standard agricultural practices for barley cultivation, providing sufficient space for robust plant growth and accurate yield measurement.
The plot area is 1 m2. Barley sowing took place on May 24, 2021, and May 25, 2022. Seeds were sown by hand in five rows at a depth of 5-7 cm (the recommended norm for this area) in plots 1.0 m long at a rate of 450 viable seeds per m2. The distance between rows was 20 cm, while the distance between plots was 50 cm. No mineral fertilizers or herbicides were used. Weeds were removed manually throughout the experiment. Artificial irrigation was not used during the entire growing season. Before sowing, seeds were treated at the rate of 200 ml of inoculant suspension (108 CFU/ml) per 1 kg of grain. The 2021 and 2022 harvests were hand-picked on August 20 and 19, respectively. Four types of biofertilizers based on highly effective strains of microorganisms were studied (Table 1).
2.5 Harvesting
For structural analysis, 25 plants in the phase of full ripeness were randomly selected. The number of stems (NSP), the number of productive stems (NPS), and the weight of 1000 grains (TW) from each variant were calculated in 3 replicates. The barley from the experimental plot was harvested by hand. The total weight of 1000 seeds was determined using a digital balance in triplicate. The total seed yield obtained from the plot was calculated in units of cwt/ha. Grain weight was measured by a random sample of 1000 grains from each treatment and expressed as the weight of 1000 grains.
2.6 Statistical analysis
All measurements were carried out in triplicate. Experimental data are presented as means and standard deviations (SD). Data were subjected to analysis of variance (ANOVA) using XL STAT. The probability of significance in ANOVA (P < 0.05) was used to indicate significant effects after biofertilizer treatment.
The main goal of the project was to study the effect of biofertilizers created in the form of consortia of microorganisms from the soils of Northern Kazakhstan on the grain yield of spring barley seeds in the conditions of the semi-arid steppe zone. To achieve the goal of the project, the seed inoculation effects of the tested biofertilizers on germination energy, seed germination, and post-embryonic development of roots and shoots of barley were studied in laboratory conditions. These parameters make it possible to assess the colonization potential of biofertilizers.
3.1 Laboratory results
According to the results of research to date, microorganisms colonize plants due to the release of organic compounds by plants and the presence of niches favorable for the development and activity of microbes. Microorganisms from the soil, atmosphere, and from the mother plant through seeds colonize three interconnected parts of plants, namely the spermosphere, rhizosphere, and phyllosphere [8]. In this project, model colonization of gnotobiotic seeds of spring barley variety “Tselinny 2005” was carried out with microorganisms from carbonate soils of southern chernozems (Table 1), and germination energy, seed germination and postembryonic development of roots and shoots of seed inoculated barley were studied (Table 2). The observed positive effect of such seed inoculation on germination energy, seed germination, and growth of barley seedlings showed that among the microorganisms of biofertilizers B1 - B4, there are bacteria and fungi that colonize seeds and stimulate the growth of seedlings. Colonization of these microorganisms occurs with the participation of germinating seeds, the exudate of which creates an environment for the growth of microorganisms [9]. Microorganisms can then colonize the roots and move into the phyllosphere to support shoot growth and development under unfavorable environmental conditions [10]. The authors found many Bacillus strains that were involved in embryo germination and seedling growth under salt-stress conditions [10].
Table 2. Effect of inoculation of barley seeds of the “Tselinny 2005” variety with biofertilizers on energy of seed germination, seed germination, and shoot development
|
Control and No. Biofertilizers |
Energy of Seed Germination, % (Second-Day) |
Seed Germination, % (Sixth-Day) |
Length of Shoots, cm (Seventh-Day) (A/B) a |
Primary Root Length, cm (Seventh-Day) (A/B) a |
Number of Lateral Roots, pcs (Seventh-Day) (A/B) a |
|
Control |
43 |
66 |
4.8±0.09 |
5.8±0.08 |
2.6 |
|
B1 |
82 (191/91) |
84 (127/27) |
6.3±0.14 (131/31) a |
7.1±0.03 (122/22) |
5.2 (200/100) |
|
B2 |
83 (193/93) |
86 (130/30) |
6.6±0.14 (138/38) a |
7.8±0.14 (134/34) |
5.0 (192/92) |
|
B3 |
57 (133/33) |
80 (121/21) |
6.0±0.06 (125/25) a |
7.1±0.08 (122/22) |
4.9 (188/88) |
|
B4 |
82 (191/91) |
85 (129/29) |
6.3±0.1 (131/31) a |
7.6±0.15 (131/31) |
5.2 (200/100) |
aA =(NF/NC) ×100% - the ratio of experimental readings to control readings in percent
aB = (NF/NC ̶ 1) ×100% - difference between experimental and control readings in percent
where, NC-measured parameter in the control variant without biofertilizer treatment
NF-measured parameter in variants with the use of biofertilizer
Variants with seeds treated with biofertilizers B1, B2, and B4 showed an increase in germination energy by 91, 93 and 91%, respectively (F=5.447, p<0.001***) and an increase in seed germination by 27, 30, and 29%, respectively (F=5.176, p<0.01**) compared to the control variant of untreated seeds. In seeds treated with B3, the increase in these parameters was more moderate and amounted to 33% and 21%, respectively. The smaller effect of biofertilizer B3 is apparently due to the different composition and number of microorganisms in the B3 consortium, which did not allow them to be effective in metabolic and colonization activity [9]. The results of seed germination given in Table 2 are in good agreement with the data of another study on the effect of treating barley seeds with isolates of Bacillus spp. from rhizosphere soil, for seed germination in laboratory conditions. The obtained germination rates of barley seeds were in the region of 85-100% [11]. Barley seeds inoculated with biofertilizers B1 - B4 in this work showed an increase in the length of shoots (F=5.447, p<0.001***), primary roots (F=5.176, p<0.01**), and the number of secondary roots, with a lesser effect of biofertilizer B3 (Table 2). Active growth in the number of secondary roots (Table 2), which is 2 times higher than the control in variants with biofertilizers B1 and B2 and lesser in other variants, shows high stimulation of the barley root system by biofertilizers in in vitro conditions when the seedlings are hungry for mineral nutrients. According to the current hypothesis, low phosphorus availability changes root architecture and promotes root branching under the influence of auxin [12]. In wild-type plants, low nitrogen content stimulated the accumulation of auxin in the lateral root primordia, which was accompanied by the appearance of additional roots [13].
The obtained stimulation of seed germination and growth of shoots, roots, and root branching in the presence of biofertilizers (Table 2) are apparently due to the synthesis of phytohormones by these microorganisms [14]. The cited authors showed that strains of methylotrophic bacteria (Methylobacterium), which in plants can produce large amounts of phytohormones (cytokinins and auxins), thrive in the tissues of spinach (Spinacia oleracea) at the cotyledon and 3-4-leaf stages, thereby promoting plant growth [14].
Significantly fewer studies have been devoted to the mechanisms of action of spermosphere microbes on plant seeds compared to the study of the mechanisms of action of rhizosphere microbes. Beneficial microorganisms that colonize seeds promote seed germination by increasing germination rate and viability [9, 14]. This is because phytohormones produced by beneficial microbial metabolism are secreted around seeds, and appropriate concentrations of phytohormones can cause changes in seed tissue morphology and increase physiological activity [15]. Seed germination and dormancy are important stages of plant growth and depend on the plant's physiology during these dormancy and germination periods [16]. The microbial community around seeds is fed by exudate metabolites from carbon deposits in the seeds. The microbial community growing in the spermosphere influences many aspects of seed biology, including germination root development, and seedling growth [17].
Thus, the results of in vitro experiments showed that microorganisms from the consortia of biofertilizers B1 – B4 can colonize not only the spermosphere and seeds but also the resulting roots of seedlings.
3.2 Results of field research
Cultivated plants grown on farms in Northern Kazakhstan inevitably face biotic and abiotic stresses of water shortage during the growing season. In the studies of this project, the agrometeorological conditions of 2021 and 2022 had a noticeable impact on the size of the spring barley harvest (Figure 1). Lack of moisture reduces transpiration and inhibits photosynthesis, which leads to metabolic disorders and damage to plant tissue [18]. Currently, plant growth-promoting rhizobacteria are being used to reduce the negative effects of drought stress on plant growth. This approach is fast-growing, environmentally friendly, and inexpensive for increasing plant productivity under abiotic stress [19]. These beneficial bacteria in the rhizosphere that promote plant growth are called Plant growth promoting rhizobacteria (PGPR). They act by increasing the availability of nutrients such as nitrogen and dissolving soil minerals such as phosphorus and potassium. They promote the production of siderophores that enhance iron assimilation [20]. They also directly stimulate plant growth by enhancing the production of phytohormones [20, 21]. PGPRs also act as antimicrobial agents that increase resistance to plant pathogens under biotic stress [13, 14]. The effectiveness of PGPR, for example, is evidenced by a high increase in barley grain yield of 149% in a plant treated with a consortium of Pseudomonas spp. and Alcaligenes faecalis [22]. PGPRs can colonize the endo- and rhizosphere of plants and cause drought resistance by producing various phytohormones, and volatile compounds, accumulating antioxidant osmolytes, producing exopolysaccharides, suppressing, or increasing the activity of stress-responsive genes, and alter root morphology, which helps withstand drought stress [23].
Microorganisms from biofertilizers B1-B4 in the abiotic and biotic extreme conditions of field experiments in this project can have the above-listed PGPR properties. To study the target issue of the project on the effect of biofertilizers on the grain yield of spring barley seeds in the conditions of a semi-arid steppe zone, two-year field tests of biofertilizers B1 - B4 were carried out (Table 3).
Table 3. Effect of biofertilizers on structural components and yield of barley seeds
|
Treatments |
NSP, pcs Number of Stems per Plant (A/B) |
NPS, pcs Number of Productive Stems (A/B) |
TW, gram Weight of 1000 Grains (A/B) |
Y, gram / m2 Productivity (A/B) |
Y, cwt/ha Productivity |
|||||
|
2021 |
2022 |
2021 |
2022 |
2021 |
2022 |
2021 |
2022 |
2021 |
2022 |
|
|
Control |
1.45±0.03 |
1.9±0.06 |
1.21±0.05 |
1.8±0.15 |
46.45±1.9 |
48.6±0.7 |
77.0±12.4 |
120.8±7.3 |
7.7 |
12.1 |
|
B1 |
2.73±0.05 188/88 p<0.05 |
2.2±0.2 116/16 p>0.05 |
2.57±0.15 212/112 p<0.05 |
2.1±0.14 117/17 p>0.05 |
53.91±1.6 116/16 p<0.05 |
51.8±0.22 107/7 p<0.05 |
129.8±22.3 169/69 p>0.05 |
249.0±4.78 206/106 p<0.05 |
13 |
24.9 |
|
С1 |
44 p<0.05 |
56 p<0.05 |
12 p<0.05 |
53 p<0.05 |
|
|
||||
|
B2 |
2.28±0.12 157/57 p<0.05 |
2.4±0.43 126/26 p>0.05 |
1.94±0.17 160/60 p<0.05 |
1.9±0.26 106/6 p>0.05 |
52.96±1.5 114/14 p<0.05 |
54.0±0.78 111/11 p<0.05 |
144.6±8.3 188/88 p<0.05 |
146.5±50.1 121/21 p>0.05 |
14.5 |
14.7 |
|
С2 |
29 p<0.05 |
30 p<0.05 |
12.5 p<0.05 |
44 p<0.05 |
|
|
||||
|
B3 |
2.12±0.12 146/46 p<0.05 |
1.8±0.17 95/0
|
1.8±0.12 149/49 p<0.05 |
1.7±0.13 94/0
|
55.8±1.15 120/20 p<0.05 |
49.9±0.22 103/3 p>0.05 |
94.7±10.6 123/23 p>0.05 |
144.5±3.9 120/20 p<0.05 |
9.5 |
14.5 |
|
С3 |
23 p<0.05 |
25 p<0.05 |
10 p<0.05 |
10 p<0.05 |
|
|
||||
|
B4 |
2.13±0.21 147/47 p<0.05 |
2.6±0.04 137/37 p<0.05 |
1.6±0.18 132/32 p<0.05 |
2.9±0.12 161/61 p<0.05 |
49.3±1.56 106/6 p>0.05 |
61.2±0.94 126/26 p<0.05 |
154.6±21.3 201/101 p<0.05 |
136±2.5 113/13 p>0.05 |
15.5 |
13.6 |
|
С4 |
42 p<0.05 |
46.5 p<0.05 |
13 p<0.05 |
51 p<0.05 |
|
|
||||
aA =(NF/NC) ×100% - the ratio of experimental readings to control readings in percent
aB = (NF/NC ̶ 1) ×100% - difference between experimental and control readings in percent
where, NC-measured parameter in the control variant without the use of biofertilizer
NF-measured parameter in variants with the use of biofertilizer
С1 – С4-average 2-year effect of biofertilizer in percent %
Nutrient availability plays a critical role in seed germination and plant growth. However, bioavailable forms of nutrients such as phosphorus and nitrogen were limited in the soil of the field experiments in this project. Pre-sowing treatment of seeds with soil microorganisms of biofertilizers B1-B4 played an important role in mitigating drought stress in plants.
According to modern concepts, in the first few days after sowing, in favorable weather conditions, seed exudates cause the migration of soil communities of microorganisms to the secreting seeds, and community of spermosphere microorganisms form around them, which ultimately contributes to the appearance of barley seedlings [9, 14]. In the conditions of field experiments biofertilizers B1 - B4 can take an active part in the process of germination of barley seedlings, and the results of laboratory and field experiments confirm their participation (Tables 2 and 3). The increase in grain yield of treated barley seeds obtained in the project is in good agreement with literature data, according to which an increase in the biotic/abiotic stress resistance of plants, and therefore productivity, is associated with plant-microbial symbiosis [24]. The maximum effect of biofertilizer B1 in increasing grain yield by 53% and its components NSP and NPS (Table 3) correlates well with the number of Bacillus spp. strains (four strains) in the tested biofertilizer (Table 1). Biofertilizer B3 contains one strain of Bacillus spp. and showed a lower percentage increase in NSP, NPS, and grain yield compared with B1 (Table 3). The 10% increase of the grain yield by B3 treatment is in good agreement with the data from another work where the increases in the grain yield of barley seeds treated by the Bacillus pumilis (M-13) and Bacillus megaterium (M-3) were equal to 7% and 8% respectively [25]. Species of the genus Bacillus more often found in scientific periodicals as phytostimulants (PGPR). When colonizing plant roots, Bacillus spp. strains exhibit direct growth-promoting mechanisms, namely siderophore production, nitrogen fixation, phytohormone production, and solubilization of phosphorus and other nutrients. Bacillus spp. strains also exhibit the indirect mechanisms of growth promotion, namely the formation of biofilms, and the production of exopolysaccharides, lytic enzymes, volatile compounds, hydrogen cyanide, etc. to promote plant growth and increase grain yield under various environmental conditions [26].
In our study, biofertilizer B3 showed a comparatively smaller impact on the germination energy of barley seeds in laboratory tests than the other biofertilizers (Table 2). This difference was quantified using statistical analysis methods, such as ANOVA, to ensure the observed effects were statistically significant. These comparisons revealed that while B3's influence was positive, it was less pronounced than that of biofertilizers B1 and B4, highlighting the variability in efficacy among the tested biofertilizers.
Inoculations of barley seeds with B1 - B4 biofertilizers in field experiments in 2021-2022 led to an average increase in the number of stems (NSP) by 44, 29, 23, and 42%, respectively, and an increase in the number of productive stems (NPS) by 56,30,25 and 47%, respectively (Table 3). The observed differences between the effects of biofertilizers on these yield components appear to be explained by the composition of taxa in the consortium of biofertilizers tested, and their biochemical and colonization properties (Tables 1 and 2). However, taxa composition was not as clearly reflected in the measurements of the TW yield component (Table 3).
Consortium of Trichoderma spp. strains from biofertilizer B4 are as effective as B1 in increasing the yield of grain and its components (Table 3), so the fungus is not inferior in properties, and in some cases superior to bacteria classified as PGPR [27]. The fungus Trichoderma cooperates with plants as a rhizosphere, epiphytic, or endophytic microorganism, never colonizing plant vascular bundles [28].
While the beneficial interactions of Trichoderma with plants are well-documented, its efficacy as a biofertilizer may exhibit discrepancies when applied across different crops or environmental conditions. Slafer and Savin [29] reported varied responses in grain yield enhancement in wheat compared to barley, suggesting species-specific interactions. Additionally, Abdelrhim et al. [30] highlighted environmental factors such as soil pH and moisture levels as critical determinants of Trichoderma's effectiveness. These findings underscore the importance of tailoring biofertilizer applications to specific agricultural contexts, acknowledging that results may not universally translate across all settings.
Interest in recent years in the Trichoderma fungi is associated with its ability to stimulate plant growth. Strains of the genus Trichoderma, like bacteria of the PGPR group, are capable of producing phytohormones, siderophores, volatile compounds, enzymes that dissolve phosphates, etc. Therefore, isolates of Trichoderma spp. that can stimulate plant growth are classified as plant growth stimulants fungi (PGPF) [31]. Regarding abiotic stresses, the use of Trichoderma spp. as a biostimulant in agriculture represents the least studied function of this group of fungi compared to its use as a biocontrol agent. Despite this, the ability of Trichoderma spp. to increase plant resistance to abiotic stresses continues to be studied [32, 33].
The grain yield increases under the influence of biofertilizers found in the present project is in good agreement with data from another study that was conducted under similar climatic conditions on a product imported from Russia [5]. The authors conducted an agroecological and bioenergy assessment of barley grain yield from the 2020 harvest, cultivated to produce concentrated feed used in the diversification of crop production. The use of a biological product during the growing season contributed to the activation of several growth, physiological, and biochemical processes of plants, which led to an increase in barley grain yield by 31%. The drug studied by the authors consisted of a liquid mixture of biologically active polyunsaturated fatty acids of the fungus Mortierella alpina and strains of the soil bacterium Azospirillum zeae VKPM B-12542 and spores of the Bacillus megaterium strain with titers of at least 1×109 CFU/ml. The TW yield component in the study was 4% [5].
However, it's noteworthy that while we observed a substantial yield boost, variations in outcomes across different studies could stem from factors such as biofertilizer composition, application rates, and crop genetic diversity. For instance, Amanullah and Khan [34] reported a more modest increase in yield, attributing the variance to different soil microbiomes' responsiveness to biofertilizers. This suggests that while biofertilizers hold promise for enhancing grain yield, their effectiveness is influenced by a complex interplay of biological and environmental factors, warranting further investigation into optimizing biofertilizer formulations and application methods for diverse agricultural contexts.
A consortium of three strains of Streptomyces spp., viz. biofertilizer B2 showed a 44% grain yield increase (Table 3), which exceeds the same parameter from the above-cited work [5]. The B2 consortium also showed NSP, NPS, and TW values comparable to these yield components of the B3 biofertilizer. The mechanisms for phytostimulants of the genus Streptomyces spp. described in the literature are closely like the mechanisms of action of PGPR and PGPF [35]. This can also explain the same percentage increase in grain yield when using biofertilizers B1 and B4 and a slightly lower percentage when using biofertilizers B2 (Table 3).
Biofertilizer B1, rich in nitrogen-fixing bacteria, significantly improved grain yield, likely due to enhanced nitrogen availability, a crucial nutrient for plant growth. Conversely, B2's lower performance might be attributed to its lesser concentration of phosphorus-solubilizing bacteria, essential for root development and energy transfer.
We believe that strains of Bacillus spp. from biofertilizers B1 and B3, strains of Streptomyces spp. from B1, B2, and B3, as well as strains of the fungus Trichoderma from B4, play a key role in barley resistance to water stress, as well as biotic stress in the experiments of this project.
Single-factor field experiments over two years showed that with the experimental dose of inoculation of barley seeds used, biofertilizers B1 and B4 are the most optimal and lead to an increase in yield by an average of 50%. The biofertilizers tested in the project can be recommended to interested agricultural enterprises as an environmentally beneficial alternative to agrochemical fertilizers and synthetic pesticides and fungicides to increase barley yields in areas subject to biotic and abiotic stress conditions.
Germination energy, germination of biofertilizer-inoculated spring barley seeds, post-embryonic development of roots and shoots measured in laboratory tests indicate effective colonization potential of at least three tested biofertilizers, viz. B1, B2 and B4.
Our analysis conclusively showed that biofertilizers B1 and B4 were responsible for a significant increase in grain yield, approximately 50%, as compared to the control group. This conclusion is directly supported by the quantitative data obtained from our field trials, where B1 and B4 treatments demonstrated a notable enhancement in both the growth rate and final yield of spring barley.
Our findings reveal that biofertilizers B1, B2, and B4 significantly outperformed B3 in promoting grain yield. For example, B1 and B4, contain strains with enhanced nitrogen-fixing capabilities or phosphorus-solubilizing bacteria, directly influencing plant growth and yield.
Microorganisms from the B3 biofertilizer consortium showed a smaller effect on the germination energy of spring barley seeds in laboratory tests and a smaller grain yield increase under conditions of abiotic and biotic stress in field tests.
According to the results of a two-year comparative study, biofertilizers B1 and B4 showed the maximum grain yield increase by about 50% and are recommended as an environmentally beneficial alternative to agrochemical fertilizers and synthetic pesticides and fungicides to increase barley grain yield in areas subject to abiotic and biotic stress conditions.
This research is funded by the Science Committee of the Ministry of Science and Higher Education of the Republic of Kazakhstan (Grant No. BR18574066).
[1] Raj, R., Shams, R., Pandey, V.K., Dash, K.K., Singh, P., Bashir, O. (2023). Barley phytochemicals and health promoting benefits: A comprehensive review. Journal of Agriculture and Food Research, 14: 100677. https://doi.org/10.1016/j.jafr.2023.100677
[2] Turuspekov, Y., Sariev, B., Chudinov, V., Sereda, G., Tokhetova, L., Ortaev, A., Tsygankov, V., Doszhanov, M., Volis, S., Abugalieva, S. (2013). Genotype×environment interaction patterns for grain yield of spring barley in different regions of Kazakhstan. Russian Journal of Genetics, 49: 196-205. https://doi.org/10.1134/S1022795413020129
[3] Khussainov, A., Ayapbergenova, A., Sarsenova, A., Aishuk, Y., Khussainova, R. (2021). Microflora, provision of ordinary chernozem with nutrients and barley productivity when inoculating the “Agrobionov” preparation. Agrivita Journal of Agricultural Science, 43(1): 13-24. https://doi.org/10.17503/agrivita.v43i1.2777
[4] ElDala, K.Z. (2023). The specific yield of grain in the Akmola region was told in the NPCC of Barayev. ElDala Media 2020.
[5] Nasiyev, B.N., Yessenguzhina, A.N. (2021). Biological technology of barley cultivation in zone 1 of Western Kazakhstan. Agrarian Science, 349(5): 71-74. https://doi.org/10.32634/0869-8155-2021-349-5-71-74
[6] Bharagava, R.N., Chandra, R., Rai, V. (2009). Isolation and characterization of aerobic bacteria capable of the degradation of synthetic and natural melanoidins from distillery effluent. World Journal of Microbiology and Biotechnology, 25: 737-744. https://doi.org/10.1007/s11274-008-9944-7
[7] ISTA. (2004). International rules for seed testing. The International Seed Testing Association, Wallisellen.
[8] Ren, X., Zhang, R., Wei, S., Luo, X., Xu, J., Shu, K. (2023). Research progress of spermosphere microorganisms. Chinese Bulletin of Botany, 58(3): 499-509. https://doi.org/10.11983/CBB22001
[9] Nelson, E.B. (2018). The seed microbiome: Origins, interactions, and impacts. Plant and Soil, 422: 7-34. https://doi.org/10.1007/s11104-017-3289-7
[10] Xu, Y., Zhang, D., Dai, L., Ding, H., Ci, D., Qin, F., Zhang, G., Zhang, Z. (2020). Influence of salt stress on growth of spermosphere bacterial communities in different peanut (Arachis hypogaea L.) cultivars. International Journal of Molecular Sciences, 21(6): 2131. https://doi.org/10.3390/ijms21062131
[11] Buntić, A., Stajković-Srbinović, O., Knežević, M., Rasulić, N., Kuzmanović, Đ., Dimitrijević-Branković, S., Delić, D. (2019). The effect of bacterial isolates from rhizosphere soils on wheat and barley seed germination. Zemljiste i Biljka, 68(2): 1-11. https://doi.org/10.5937/zembilj1902001b
[12] López-Bucio, J., Hernández-Abreu, E., Sánchez-Calderón, L., Pérez-Torres, A., Rampey, R.A., Bartel, B., Herrera-Estrella, L. (2005). An auxin transport independent pathway is involved in phosphate stress-induced root architectural alterations in Arabidopsis. Identification of BIG as a mediator of auxin in pericycle cell activation. Plant Physiology, 137(2): 681-691. https://doi.org/10.1104/pp.104.049577
[13] Ma, W., Li, J., Qu, B., He, X., Zhao, X., Li, B., Fu, X., Tong, Y. (2014). Auxin biosynthetic gene TAR2 is involved in low nitrogen-mediated reprogramming of root architecture in arabidopsis. Plant Journal, 78(1): 70-79. https://doi.org/10.1111/tpj.12448
[14] Lopez-Velasco, G., Carder, P.A., Welbaum, G.E., Ponder, M.A. (2013). Diversity of the spinach (Spinacia oleracea) spermosphere and phyllosphere bacterial communities. FEMS Microbiology Letters, 346(2): 146-154. https://doi.org/10.1111/1574-6968.12216
[15] Dodd, I.C., Zinovkina, N.Y., Safronova, V.I., Belimov, A.A. (2010). Rhizobacterial mediation of plant hormone status. Annals of Applied Biology, 157(3): 361-379. https://doi.org/10.1111/j.1744-7348.2010.00439.x
[16] Shu, K., Liu, X.D., Xie, Q., He, Z.H. (2016). Two faces of on seed: Hormonal regulation of dormancy and germination. Molecular Plant, 9(1): 34-45. http://dx.doi.org/10.1016/j.molp.2015.08.010
[17] Aziz, U., Rehmani, M.S., Wang, L., Luo, X.F., Xian, B.S., Wei, S.W., Wang, G.D., Shu, K. (2021). Toward a molecular understanding of rhizosphere, phyllosphere, and spermosphere interactions in plant growth and stress response. Critical Reviews in Plant Sciences, 40(6): 479-500. https://doi.org/10.1080/07352689.2022.2031728
[18] Gul, F., Khan, I.U., Rutherford, S., Dai, Z.C., Li, G., Du, D.L. (2023). Plant growth promoting rhizobacteria and biochar production from Parthenium hysterophorus enhance seed germination and productivity in barley under drought stress. Frontiers in Plant Science, 14: 1175097. https://doi.org/doi:10.3389/fpls.2023.1175097
[19] Akhtar, N., Ilyas, N., Mashwani, Z.-U.-R., Hayat, R., Yasmin, H., Noureldeen, A., Ahmad, P. (2021). Synergistic effects of plant growth promoting rhizobacteria and silicon dioxide nanoparticles for amelioration of drought stress in wheat. Plant Physiology and Biochemistry, 166: 160-176. https://doi.org/10.1016/j.plaphy.2021.05.039
[20] Bhat, M.A., Rasool, R., Ramzan, S. (2019). Plant growth promoting rhizobacteria (PGPR) for sustainable and eco-friendly agriculture. Acta Scientific Agriculture, 3(1): 23-25.
[21] Khan, N., Bano, A., Ali, S., Babar, Md.A. (2020). Crosstalk amongst phytohormones from planta and PGPR under biotic and abiotic stresses. Plant Growth Regulation, 90: 189-203. https://doi.org/10.1007/s10725-020-00571-x
[22] Divyanshu, K., Yadav, M., Upadhyay, R.S. (2022). Effectiveness of pseudomonas and alcaligenes sp. on the yield and lignin deposition of barley hordeum vulgare L. crop. Bioscience Biotechnology Research Communications, 15(3): 466-474. https://doi.org/10.21786/bbrc/15.3.14
[23] Kumari, S., Vaishnav, A., Jain, S., Varma, A., Choudhary, D.K. (2015). Bacterial-mediated induction of systemic tolerance to salinity with expression of stress alleviating enzymes in soybean (Glycine max L. Merrill). Plant Growth Regulation, 34: 558-573. https://doi.org/10.1007/s00344-015-9490-0
[24] Mastouri, F., Björkman, T., Harman, G.E. (2010). Seed treatment with Trichoderma harzianum alleviates biotic, abiotic, and physiological stresses in germinating seeds and seedlings. Phytopathology, 100(11): 1213-1221. https://doi.org/10.1094/PHYTO-03-10-0091
[25] Çakmakçi, R., Kantar, F., Sahin, F. (2001). Effect of N2-fixing bacterial inoculations on yield of sugar beet and barley. Journal of Plant Nutrition and Soil Science, 164(5): 527-531. https://doi.org/10.1002/1522-2624(200110)164:5<527::AID-JPLN527>3.0.CO;2-1
[26] Tsotetsi, T., Nephali, L., Malebe, M., Tugizimana, F. (2022). Bacillus for plant growth promotion and stress resilience: What have we learned? Plants, 11(19): 2482. https://doi.org/10.3390/plants11192482
[27] Khan, M.R., Mohiddin, F.A. (2018). Trichoderma: Its multifarious utility in crop improvement. In: Prasad, R., Gill, S.S., Tuteja, N. (eds) Crop Improvement through Microbial Biotechnology, pp. 263-291. https://doi.org/10.1016/B978-0-444-63987-5.00013-X
[28] Poveda, J., Eugui, D., Abril-Urias, P. (2020). Could Trichoderma be a plant pathogen? Successful root colonization. In: Sharma, A., Sharma, P. (eds) Trichoderma. Rhizosphere Biology, Singapore, pp. 35-59. https://doi.org/10.1007/978-981-15-3321-1_3
[29] Slafer, G.A., Savin, R. (2023). Comparative performance of barley and wheat across a wide range of yielding conditions. Does barley outyield wheat consistently in low-yielding conditions? European Journal of Agronomy, 143. https://doi.org/10.1016/j.eja.2022.126689
[30] Abdelrhim, A.S., Hemeda, N.F., Mwaheb, M.A., Omar, M.O.A., Dawood, M.F.A. (2024). The role of Trichoderma koningii and Trichoderma harzianum in mitigating the combined stresses motivated by Sclerotiniasclerotiorum and salinity in common bean (Phaseolusvulgaris). Plant Stress, 11. https://doi.org/10.1016/j.stress.2024.100370
[31] El Enshasy, H.A., Ambehabati, K.K., El Baz, A.F., Ramchuran, S., Sayyed, R.Z., Amalin, D., Dailin, D.J., Hanapi, S.Z. (2020). Trichoderma: biocontrol agents for promoting plant growth and soil health. Agriculturally Important Fungi for Sustainable Agriculture, 239-259. https://doi.org/10.1007/978-3-030-48474-3_8
[32] Zaidi, N.W., Dar, M.H., Singh, S., Singh, U.S. (2014). Trichoderma species as abiotic stress relievers in plants. In: Biotechnology and biology of Trichoderma, pp. 515-525. https://doi.org/10.1016/B978-0-444-59576-8.00038-2
[33] Hidangmayum, A., Dwivedi, P. (2018). Plant responses to Trichoderma spp. and their tolerance to abiotic stresses: A review. Journal of Pharmacognosy and Phytochemistry, 7(1): 758-766.
[34] Amanullah, A., Khan, U. (2023). Advancing sustainable agriculture with beneficial microbes: Enhancing crop growth and yield for food security and human health. Advances in Modern Agriculture, 4(2). https://doi.org/10.54517/ama.v4i2.2426
[35] Silva, G.C., Kitano, I.T., Ribeiro, I.A.F., Lacava, P.T. (2022). The potential use of actinomycetes as microbial inoculants and biopesticides in agriculture. Frontiers in Soil Science, 2: 833181. https://doi.org/10.3389/fsoil.2022.833181